| Dinosaurus Rentang waktu: (kemungkinan catatan Trias Tengah)
| |||
|---|---|---|---|
 Herrerasaurus ischigualastensis (dinosaurus basal karnivora)    | |||
| Klasifikasi ilmiah | |||
| Kerajaan: | Animalia | ||
| Filum: | Chordata | ||
| Klad: | Sauropsida | ||
| Klad: | Archosauria | ||
| Klad: | Avemetatarsalia | ||
| Klad: | Ornithodira | ||
| Klad: | Dinosauromorpha | ||
| Klad: | Dinosauriformes | ||
| Klad: | Dracohors | ||
| Klad: | Dinosauria Owen, 1842 | ||
| Kelompok utama | |||
| |||



Dinosaurus adalah kelompok reptil yang beragam[note 1] dari klad Dinosauria. Dinosaurus pertama kali muncul selama periode Trias, antara 243 dan 233,23 juta tahun lalu (jtl), meskipun asal-usul dan waktu yang tepat dari evolusi dinosaurus masih menjadi subjek penelitian aktif. Dinosaurus menjadi vertebrata darat yang dominan setelah peristiwa kepunahan Trias–Jura 201,3 jtl dan dominasi mereka berlanjut sepanjang periode Jura dan Kapur. Catatan fosil menunjukkan bahwa burung adalah dinosaurus berbulu, yang berevolusi dari theropoda awal selama kala Jura Akhir, dan merupakan satu-satunya garis keturunan dinosaurus yang diketahui selamat dari peristiwa kepunahan Kapur–Paleogen sekitar 66 jtl. Oleh karena itu, dinosaurus dapat dibagi menjadi dinosaurus burung (avian) dan dinosaurus non-burung yang telah punah, yaitu semua dinosaurus selain burung.
Dinosaurus sangat bervariasi dari sudut pandang taksonomi, morfologi, dan ekologi. Burung, dengan lebih dari 11.000 spesies yang hidup, merupakan salah satu kelompok vertebrata yang paling beragam. Menggunakan bukti fosil, paleontolog telah mengidentifikasi lebih dari 900 genera berbeda dan lebih dari 1.000 spesies dinosaurus non-avian yang berbeda. Dinosaurus terwakili di setiap benua baik oleh spesies ekstan (burung) maupun sisa-sisa fosil. Sepanjang sebagian besar abad ke-20, sebelum burung diakui sebagai dinosaurus, sebagian besar komunitas ilmiah meyakini dinosaurus sebagai hewan yang lamban dan berdarah dingin. Namun, sebagian besar penelitian yang dilakukan sejak tahun 1970-an telah mengindikasikan bahwa dinosaurus adalah hewan yang aktif dengan metabolisme tinggi dan berbagai adaptasi untuk interaksi sosial. Beberapa bersifat herbivora, sementara yang lain karnivora. Bukti menunjukkan bahwa semua dinosaurus bertelur, dan bahwa pembuatan sarang adalah sifat yang dimiliki oleh banyak dinosaurus, baik avian maupun non-avian.
Meskipun nenek moyang dinosaurus bersifat bipedal, banyak kelompok yang telah punah mencakup spesies kuadrupedal, dan beberapa mampu beralih di antara kedua postur ini. Struktur peragaan yang rumit seperti tanduk atau jambul umum ditemukan di semua kelompok dinosaurus, dan beberapa kelompok yang telah punah mengembangkan modifikasi rangka seperti baju zirah tulang dan duri. Sementara garis keturunan avian dinosaurus yang bertahan hingga masa kini (burung) umumnya berukuran kecil karena kendala kemampuan terbang, banyak dinosaurus non-avian bertubuh besar—dinosaurus sauropoda terbesar diperkirakan mencapai panjang 397 meter (1.302 kaki) dan tinggi 18 m (59 ft) serta merupakan hewan darat terbesar sepanjang masa.
Fosil dinosaurus pertama dikenali pada awal abad ke-19, dengan nama "dinosaur" (berarti "kadal yang mengerikan") dicetuskan oleh Sir Richard Owen pada tahun 1842 untuk merujuk pada "kadal fosil raksasa" ini. Sejak saat itu, kerangka fosil dinosaurus yang dipajang telah menjadi atraksi utama di museum-museum di seluruh dunia, dan dinosaurus telah menjadi bagian abadi dari budaya populer. Ukuran besar dari beberapa dinosaurus, serta sifat mereka yang tampak mengerikan dan fantastis, telah memastikan kemunculan rutin mereka dalam buku dan film terlaris, seperti waralaba Jurassic Park. Antusiasme publik yang terus-menerus terhadap hewan-hewan ini telah menghasilkan pendanaan yang signifikan bagi penelitian dinosaurus, dan penemuan-penemuan baru secara rutin diliput oleh media.
Definisi
suntingBerdasarkan tata nama filogenetik, dinosaurus biasanya didefinisikan sebagai kelompok yang terdiri dari leluhur bersama terkini (most recent common ancestor/MRCA) dari Triceratops dan burung modern (Neornithes), serta semua keturunannya.[7] Disarankan juga bahwa Dinosauria didefinisikan sehubungan dengan MRCA dari Megalosaurus dan Iguanodon, karena keduanya adalah dua dari tiga genera yang dikutip oleh Richard Owen ketika ia mengenali Dinosauria.[8] Kedua definisi tersebut mencakup genera umum yang sama: Dinosauria = Ornithischia + Saurischia. Ini mencakup kelompok-kelompok utama seperti ankylosauria (kuadruped herbivora berbaju zirah), stegosauria (kuadruped herbivora berlempeng), ceratopsia (herbivora bipedal atau kuadrupedal dengan jumbai leher), pachycephalosauria (herbivora bipedal dengan tengkorak tebal), ornithopoda (herbivora bipedal atau kuadrupedal termasuk "paruh bebek"), theropoda (sebagian besar karnivora bipedal dan burung), serta sauropodomorpha (sebagian besar kuadruped herbivora besar dengan leher dan ekor panjang).[9]
Burung adalah satu-satunya dinosaurus yang masih bertahan hidup. Dalam taksonomi tradisional, burung dianggap sebagai kelas terpisah yang berevolusi dari dinosaurus. Namun, sebagian besar paleontolog kontemporer menolak gaya klasifikasi tradisional yang didasarkan pada kesamaan anatomi, dan lebih memilih taksonomi filogenetik berdasarkan garis keturunan yang dideduksi, di mana setiap kelompok didefinisikan sebagai semua keturunan dari genus pendiri tertentu.[10] Burung termasuk dalam subkelompok dinosaurus Maniraptora, yang merupakan coelurosauria, yang merupakan theropoda, yang merupakan saurischia.[11]
Penelitian oleh Matthew G. Baron, David B. Norman, dan Paul M. Barrett pada tahun 2017 menyarankan revisi radikal terhadap sistematika dinosaurus. Analisis filogenetik oleh Baron et al. menemukan bahwa Ornithischia lebih dekat dengan Theropoda daripada Sauropodomorpha, berlawanan dengan penyatuan tradisional theropoda dengan sauropodomorpha. Hal ini akan menyebabkan sauropoda dan kerabatnya berada di luar dinosaurus tradisional, sehingga mereka mendefinisikan ulang Dinosauria sebagai leluhur bersama terakhir dari Triceratops horridus, Passer domesticus dan Diplodocus carnegii, serta semua keturunannya, untuk memastikan bahwa sauropoda dan kerabatnya tetap termasuk sebagai dinosaurus. Mereka juga membangkitkan kembali klad Ornithoscelida untuk merujuk pada kelompok yang berisi Ornithischia dan Theropoda.[12][13]
Deskripsi umum
sunting
Menggunakan salah satu definisi di atas, dinosaurus secara umum dapat dideskripsikan sebagai archosaur dengan tungkai belakang yang tegak di bawah tubuh.[14] Hewan prasejarah lainnya, termasuk pterosaurus, mosasaurus, ichthyosaurus, plesiosaurus, dan Dimetrodon, meskipun sering dianggap secara populer sebagai dinosaurus, secara taksonomi tidak diklasifikasikan sebagai dinosaurus. Pterosaurus berkerabat jauh dengan dinosaurus, menjadi anggota klad Ornithodira. Kelompok lain yang disebutkan, seperti halnya dinosaurus dan pterosaurus, adalah anggota Sauropsida (klad reptil dan burung), kecuali Dimetrodon (yang merupakan sinapsida). Tak satu pun dari mereka memiliki postur tungkai belakang tegak yang menjadi karakteristik dinosaurus sejati.[15]
Dinosaurus adalah vertebrata darat yang dominan pada Era Mesozoikum, terutama periode Jura dan Kapur. Kelompok hewan lain terbatas dalam ukuran dan relung; mamalia, misalnya, jarang melebihi ukuran kucing domestik dan umumnya merupakan karnivora seukuran hewan pengerat yang memangsa buruan kecil.[16] Dinosaurus selalu diakui sebagai kelompok yang sangat bervariasi: lebih dari 900 genus dinosaurus non-avian telah diidentifikasi secara meyakinkan (2018) dengan 1.124 spesies (2016). Estimasi menempatkan jumlah total genus dinosaurus yang terawetkan dalam catatan fosil pada angka 1.850, dengan hampir 75% masih belum ditemukan,[17][18][19] dan jumlah yang pernah ada (di dalam atau di luar catatan fosil) mencapai 3.400.[20] Sebuah estimasi tahun 2016 menempatkan jumlah spesies dinosaurus yang hidup di Mesozoikum pada kisaran 1.543–2.468,[21][22] dibandingkan dengan jumlah burung modern (dinosaurus avian) sebanyak 10.806 spesies.[23]
Dinosaurus yang telah punah, serta burung modern, mencakup genera yang bersifat herbivora dan yang lain karnivora, termasuk pemakan biji, pemakan ikan, insektivora, dan omnivora. Meskipun dinosaurus secara leluhur bersifat bipedal (seperti halnya semua burung modern), beberapa berevolusi menjadi kuadruped, dan yang lain, seperti Anchisaurus dan Iguanodon, dapat berjalan dengan sama mudahnya menggunakan dua atau empat kaki. Modifikasi kranial seperti tanduk dan jambul adalah ciri umum dinosaurus, dan beberapa spesies yang telah punah memiliki baju zirah tulang. Meskipun genera yang paling terkenal luar biasa karena ukurannya yang besar, banyak dinosaurus Mesozoikum berukuran seukuran manusia atau lebih kecil, dan burung modern umumnya berukuran kecil. Dinosaurus saat ini menghuni setiap benua, dan fosil menunjukkan bahwa mereka telah mencapai penyebaran global paling lambat pada kala Jura Awal.[24] Burung modern menghuni sebagian besar habitat yang tersedia, dari terestrial hingga laut, dan terdapat bukti bahwa beberapa dinosaurus non-avian (seperti Microraptor) dapat terbang atau setidaknya meluncur, dan yang lain, seperti spinosauridae, memiliki kebiasaan semiakuatik.[25]
Ciri-ciri anatomi pembeda
suntingMeskipun penemuan terbaru membuat penyajian daftar fitur pembeda yang disepakati secara universal menjadi lebih sulit, hampir semua dinosaurus yang ditemukan sejauh ini berbagi modifikasi tertentu pada kerangka archosauria leluhur, atau secara jelas merupakan keturunan dari dinosaurus yang lebih tua yang menunjukkan modifikasi tersebut. Walaupun beberapa kelompok dinosaurus kemudian menampilkan versi modifikasi lebih lanjut dari sifat-sifat ini, ciri-ciri tersebut dianggap khas untuk Dinosauria; dinosaurus paling awal memilikinya dan mewariskannya kepada keturunan mereka. Modifikasi semacam itu, yang berasal dari leluhur bersama terkini dari kelompok taksonomi tertentu, disebut sebagai sinapomorfi dari kelompok tersebut.[26]
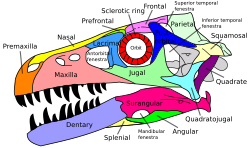
Penilaian mendetail mengenai hubungan antar-archosauria oleh Sterling Nesbitt[27] mengonfirmasi atau menemukan dua belas sinapomorfi tegas berikut, beberapa di antaranya telah diketahui sebelumnya:
- Pada tengkorak, terdapat fossa supratemporal (cekungan) di depan fenestra supratemporal, bukaan utama di bagian belakang atap tengkorak
- Epipofisis, prosesus (unjuran) yang mengarah miring ke belakang pada sudut atas bagian belakang vertebra leher anterior (depan) di belakang atlas dan axis, dua vertebra leher pertama
- Puncak dari krista deltopektoralis (tonjolan tempat otot deltopektoral melekat) terletak pada atau lebih dari 30% ke bawah sepanjang humerus (tulang lengan atas)
- Radius, tulang lengan bawah, lebih pendek dari 80% panjang humerus
- Trokanter keempat (tonjolan tempat otot caudofemoralis melekat pada batang belakang bagian dalam) pada femur (tulang paha) merupakan flens yang tajam
- Trokanter keempat asimetris, dengan tepi distal (bawah) membentuk sudut yang lebih curam terhadap batang tulang
- Pada astragalus dan kalkaneus, tulang pergelangan kaki atas, faset artikular proksimal (permukaan penghubung atas) untuk fibula menempati kurang dari 30% lebar transversal elemen tersebut
- Exoccipital (tulang di bagian belakang tengkorak) tidak bertemu di sepanjang garis tengah pada dasar rongga endokranial, ruang bagian dalam tempurung otak
- Pada panggul, permukaan artikular proksimal ischium dengan ilium dan pubis dipisahkan oleh permukaan cekung yang besar (pada sisi atas ischium, bagian dari sendi panggul yang terbuka terletak di antara kontak dengan tulang pubis dan ilium)
- Krista cnemial pada tibia (bagian yang menonjol dari permukaan atas tulang kering) melengkung secara anterolateral (melengkung ke depan dan sisi luar)
- Punggungan berorientasi proksimodistal (vertikal) yang jelas terdapat pada permukaan posterior ujung distal tibia (permukaan belakang ujung bawah tulang kering)
- Permukaan artikular cekung untuk fibula pada kalkaneus (permukaan atas kalkaneus, tempat bersentuhan dengan fibula, memiliki profil berongga)
Nesbitt menemukan sejumlah potensi sinapomorfi lanjutan dan mengabaikan sejumlah sinapomorfi yang disarankan sebelumnya. Beberapa di antaranya juga terdapat pada silesaurida, yang oleh Nesbitt ditempatkan sebagai kelompok saudari bagi Dinosauria, termasuk trokanter anterior yang besar, metatarsal II dan IV dengan panjang yang hampir sama, kontak yang berkurang antara ischium dan pubis, adanya krista cnemial pada tibia dan prosesus asenden pada astragalus, serta banyak lainnya.[7]

Berbagai fitur kerangka lainnya juga dimiliki bersama oleh dinosaurus. Namun, karena fitur-fitur tersebut umum ditemukan pada kelompok archosauria lain atau tidak ada pada semua dinosaurus awal, fitur-fitur ini bukanlah sinapomorfi. Sebagai contoh, sebagai diapsida, dinosaurus secara leluhur memiliki dua pasang fenestra infratemporal (bukaan pada tengkorak di belakang mata), dan sebagai anggota kelompok diapsida Archosauria, memiliki bukaan tambahan pada moncong dan rahang bawah.[28] Selain itu, beberapa karakteristik yang pernah dianggap sebagai sinapomorfi kini diketahui telah muncul sebelum dinosaurus, atau tidak ada pada dinosaurus paling awal dan berevolusi secara independen oleh kelompok dinosaurus yang berbeda. Ini termasuk skapula, atau tulang belikat, yang memanjang; sakrum yang terdiri dari tiga atau lebih vertebra yang menyatu (tiga ditemukan pada beberapa archosauria lain, tetapi hanya dua yang ditemukan pada Herrerasaurus);[7] dan asetabulum, atau soket panggul, yang berlubang, dengan lubang di tengah permukaan bagian dalamnya (tertutup pada Saturnalia tupiniquim, misalnya).[29][30] Kesulitan lain dalam menentukan fitur khas dinosaurus adalah bahwa dinosaurus awal dan archosauria lain dari kala Trias Akhir sering kali kurang dikenal dan serupa dalam banyak hal; hewan-hewan ini terkadang salah diidentifikasi dalam literatur.[31]
Dinosaurus berdiri dengan tungkai belakang tegak dengan cara yang mirip dengan sebagian besar mamalia modern, tetapi berbeda dari sebagian besar reptil lain, yang tungkainya merentang ke samping.[32] Postur ini disebabkan oleh perkembangan ceruk yang menghadap ke samping pada panggul (biasanya berupa soket terbuka) dan kepala tulang yang jelas menghadap ke dalam pada femur.[33] Postur tegak mereka memungkinkan dinosaurus awal untuk bernapas dengan mudah saat bergerak, yang kemungkinan memungkinkan stamina dan tingkat aktivitas yang melampaui reptil yang "merayap".[34] Tungkai tegak mungkin juga membantu mendukung evolusi ukuran tubuh yang besar dengan mengurangi tekanan tekuk pada tungkai.[35] Beberapa archosauria non-dinosaurus, termasuk rauisuchia, juga memiliki tungkai tegak tetapi mencapainya dengan konfigurasi sendi panggul "tegak-pilar", di mana alih-alih memiliki tonjolan dari femur yang masuk ke dalam soket pada panggul, tulang panggul atas diputar untuk membentuk rak yang menggantung.[35]
Sejarah penelitian
suntingSejarah pra-ilmiah
suntingFosil dinosaurus telah dikenal selama ribuan tahun, meskipun sifat aslinya tidak dikenali. Orang Tiongkok menganggapnya sebagai tulang naga dan mendokumentasikannya demikian. Sebagai contoh, Huayang Guo Zhi (華陽國志), sebuah catatan wilayah yang disusun oleh Chang Qu (常璩) selama Dinasti Jin Barat (265–316), melaporkan penemuan tulang naga di Wucheng, Provinsi Sichuan.[36] Penduduk desa di Tiongkok tengah telah lama menggali "tulang naga" yang terfosilisasi untuk digunakan dalam obat-obatan tradisional.[37] Di Eropa, fosil dinosaurus umumnya diyakini sebagai sisa-sisa raksasa dan makhluk alkitabiah lainnya.[38]
Penelitian awal dinosaurus
sunting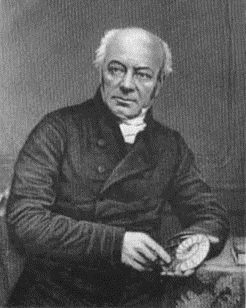
Deskripsi ilmiah mengenai apa yang sekarang dikenal sebagai tulang dinosaurus pertama kali muncul pada akhir abad ke-17 di Inggris. Bagian dari tulang, yang sekarang diketahui sebagai tulang paha (femur) dari Megalosaurus,[39] ditemukan dari tambang batu gamping di Cornwell dekat Chipping Norton, Oxfordshire, pada tahun 1676. Fragmen tersebut dikirim ke Robert Plot, Profesor Kimia di Universitas Oxford dan kurator pertama Museum Ashmolean, yang menerbitkan deskripsinya dalam The Natural History of Oxford-shire (1677).[40] Ia dengan tepat mengidentifikasi tulang tersebut sebagai ujung bawah dari femur hewan besar, dan menyadari bahwa tulang itu terlalu besar untuk menjadi milik spesies yang dikenal. Oleh karena itu, ia menyimpulkan bahwa tulang itu adalah femur manusia raksasa, mungkin seorang Titan atau jenis raksasa lain yang diceritakan dalam legenda.[41][42] Edward Lhuyd, teman dari Sir Isaac Newton, menerbitkan Lithophylacii Britannici ichnographia (1699), perlakuan ilmiah pertama terhadap apa yang sekarang dikenal sebagai dinosaurus. Di dalamnya ia mendeskripsikan dan menamai sebuah gigi sauropoda, "Rutellum impicatum",[43][44] yang telah ditemukan di Caswell, dekat Witney, Oxfordshire.[45]

Antara tahun 1815 dan 1824, Pdt. William Buckland, Dosen Geologi pertama di Universitas Oxford, mengumpulkan lebih banyak tulang terfosilisasi dari Megalosaurus dan menjadi orang pertama yang mendeskripsikan dinosaurus non-avian dalam sebuah jurnal ilmiah.[39][46] Genus dinosaurus non-avian kedua yang diidentifikasi, Iguanodon, konon ditemukan pada tahun 1822 oleh Mary Ann Mantell, istri geolog Inggris Gideon Mantell, meskipun hal ini diperdebatkan dan beberapa sejarawan mengatakan Gideon telah memperoleh sisa-sisa tersebut bertahun-tahun sebelumnya. Gideon Mantell mengenali kesamaan antara fosilnya dan tulang iguana modern serta menerbitkan temuannya pada tahun 1825.[47][48]
Penelitian tentang "kadal fosil raksasa" ini segera menjadi perhatian besar bagi para ilmuwan Eropa dan Amerika, dan pada tahun 1842 paleontolog Inggris Sir Richard Owen mencetuskan istilah "dinosaur", menggunakannya untuk merujuk pada "suku atau sub-ordo Reptil Sauria yang berbeda" yang kemudian dikenal di Inggris dan di seluruh dunia.[49][50][51][52][53] Istilah ini berasal dari bahasa Yunani Kuno δεινός (deinos), artinya "mengerikan, kuat, atau sangat hebat", dan σαῦρος (sauros), artinya "kadal atau reptil".[52][54] Meskipun nama taksonomi tersebut sering ditafsirkan sebagai rujukan pada gigi, cakar, dan karakteristik menakutkan lainnya dari dinosaurus, Owen bermaksud agar nama itu juga membangkitkan kesan ukuran dan keagungan mereka.[55] Owen menyadari bahwa sisa-sisa yang telah ditemukan sejauh ini, Iguanodon, Megalosaurus, dan Hylaeosaurus, memiliki ciri-ciri khas yang sama, sehingga memutuskan untuk menyajikannya sebagai kelompok taksonomi yang berbeda. Sebagaimana diklarifikasi oleh geolog dan sejarawan Inggris Hugh Torrens, Owen telah memberikan presentasi tentang reptil fosil kepada British Association for the Advancement of Science pada tahun 1841, tetapi laporan pada saat itu menunjukkan bahwa Owen tidak menyebutkan kata "dinosaur", atau mengakui dinosaurus sebagai kelompok reptil yang berbeda dalam pidatonya. Ia memperkenalkan Dinosauria hanya dalam versi teks revisi dari pidatonya yang diterbitkan pada bulan April 1842.[49][50] Dengan dukungan dari Pangeran Albert, suami Ratu Victoria, Owen mendirikan Museum Sejarah Alam, London, untuk memamerkan koleksi nasional fosil dinosaurus serta pameran biologi dan geologi lainnya.[56]
Penemuan di Amerika Utara
sunting

Pada tahun 1858, William Parker Foulke menemukan dinosaurus Amerika pertama yang diketahui, di lubang galian tanah napal di kota kecil Haddonfield, New Jersey. (Meskipun fosil telah ditemukan sebelumnya, sifat aslinya belum dipahami dengan benar.) Makhluk itu dinamai Hadrosaurus foulkii. Ini adalah temuan yang sangat penting: Hadrosaurus adalah salah satu kerangka dinosaurus hampir lengkap pertama yang ditemukan (yang pertama ditemukan pada tahun 1834, di Maidstone, Inggris), dan itu jelas merupakan makhluk bipedal. Ini adalah penemuan revolusioner karena, sampai saat itu, sebagian besar ilmuwan percaya dinosaurus berjalan dengan empat kaki, seperti kadal lainnya. Penemuan Foulke memicu gelombang minat terhadap dinosaurus di Amerika Serikat, yang dikenal sebagai mania dinosaurus.[57]
Mania dinosaurus dicontohkan oleh persaingan sengit antara Edward Drinker Cope dan Othniel Charles Marsh, yang keduanya berlomba menjadi yang pertama menemukan dinosaurus baru dalam apa yang kemudian dikenal sebagai Perang Tulang. Pertarungan antara kedua ilmuwan ini berlangsung selama lebih dari 30 tahun, berakhir pada tahun 1897 ketika Cope meninggal setelah menghabiskan seluruh kekayaannya untuk perburuan dinosaurus. Banyak spesimen dinosaurus berharga yang rusak atau hancur karena metode kasar pasangan tersebut: misalnya, penggali mereka sering menggunakan dinamit untuk menggali tulang. Paleontolog modern akan menganggap metode seperti itu kasar dan tidak dapat diterima, karena peledakan dengan mudah menghancurkan bukti fosil dan stratigrafi. Meskipun metode mereka tidak halus, kontribusi Cope dan Marsh bagi paleontologi sangat luas: Marsh menggali 86 spesies baru dinosaurus dan Cope menemukan 56, dengan total 142 spesies baru. Koleksi Cope sekarang berada di Museum Sejarah Alam Amerika di Kota New York, sementara koleksi Marsh berada di Museum Sejarah Alam Peabody di Universitas Yale.[58]
"Renaisans dinosaurus" dan seterusnya
suntingPerang Dunia II menyebabkan jeda dalam penelitian paleontologi; setelah perang, perhatian penelitian juga semakin dialihkan ke fosil mamalia daripada dinosaurus, yang dipandang lamban dan berdarah dingin.[59][60] Namun, pada akhir tahun 1960-an, bidang penelitian dinosaurus mengalami lonjakan aktivitas yang masih berlangsung hingga kini.[61] Beberapa studi penting memicu aktivitas ini. Pertama, John Ostrom menemukan theropoda dromaeosaurid yang mirip burung, Deinonychus, dan mendeskripsikannya pada tahun 1969. Anatominya mengindikasikan bahwa ia adalah predator aktif yang kemungkinan berdarah panas, sangat kontras dengan citra dinosaurus yang berlaku saat itu.[59] Secara bersamaan, Robert T. Bakker menerbitkan serangkaian studi yang juga berpendapat mengenai gaya hidup aktif pada dinosaurus berdasarkan bukti anatomi dan ekologi (lihat § Fisiologi),[62][63] yang kemudian diringkas dalam bukunya tahun 1986 The Dinosaur Heresies.[64]

Pengungkapan baru didukung oleh peningkatan penemuan dinosaurus. Penemuan dinosaurus baru yang besar telah dilakukan oleh para paleontolog yang bekerja di wilayah yang sebelumnya belum dieksplorasi, termasuk India, Amerika Selatan, Madagaskar, Antartika, dan yang paling signifikan Tiongkok. Di seluruh kelompok theropoda, sauropodomorpha, dan ornithischia, jumlah genera yang dinamai mulai meningkat secara eksponensial pada tahun 1990-an.[17] Pada tahun 2008, lebih dari 30 spesies dinosaurus baru dinamai setiap tahunnya.[65] Setidaknya sauropodomorpha mengalami peningkatan lebih lanjut dalam jumlah spesies yang dinamai pada tahun 2010-an, dengan rata-rata 9,3 spesies baru dinamai setiap tahun antara 2009 dan 2020. Sebagai konsekuensinya, lebih banyak sauropodomorpha yang dinamai antara tahun 1990 dan 2020 dibandingkan gabungan tahun-tahun sebelumnya.[66] Lokasi-lokasi baru ini juga mengarah pada peningkatan kualitas spesimen secara keseluruhan, dengan spesies baru yang semakin banyak dinamai bukan berdasarkan fosil yang serpihan melainkan pada kerangka yang lebih lengkap, terkadang dari beberapa individu. Spesimen yang lebih baik juga menyebabkan spesies baru lebih jarang dibatalkan validitasnya.[65] Lokasi Asia telah menghasilkan spesimen theropoda paling lengkap,[67] sementara lokasi Amerika Utara telah menghasilkan spesimen sauropodomorpha paling lengkap.[66]
Sebelum renaisans dinosaurus, dinosaurus sebagian besar diklasifikasikan menggunakan sistem berbasis peringkat tradisional dari taksonomi Linnaean. Renaisans ini juga disertai dengan penerapan kladistik yang makin luas, sebuah metode klasifikasi yang lebih objektif berdasarkan garis keturunan dan ciri-ciri bersama, yang terbukti sangat berguna dalam studi sistematika dan evolusi dinosaurus. Analisis kladistik, di antara teknik-teknik lainnya, membantu mengompensasi catatan fosil yang sering kali tidak lengkap dan terfragmentasi.[68][69] Buku referensi yang merangkum keadaan penelitian dinosaurus, seperti The Dinosauria karya David B. Weishampel dan rekan-rekannya, membuat pengetahuan lebih mudah diakses[70] dan memacu minat lebih lanjut dalam penelitian dinosaurus. Rilis edisi pertama dan kedua dari The Dinosauria pada tahun 1990 dan 2004, serta makalah tinjauan oleh Paul Sereno pada tahun 1998, disertai dengan peningkatan jumlah pohon filogenetika yang diterbitkan untuk dinosaurus.[71]
Pengawetan jaringan lunak dan molekuler
sunting
Fosil dinosaurus tidak terbatas pada tulang, tetapi juga mencakup cetakan atau sisa-sisa terimineralisasi dari penutup kulit, organ, dan jaringan lainnya. Di antaranya, penutup kulit yang berbasis protein keratin adalah yang paling mudah terawetkan karena struktur molekulnya yang terhubung silang dan hidrofobik.[72] Fosil penutup kulit berbasis keratin atau penutup kulit bertulang diketahui dari sebagian besar kelompok utama dinosaurus. Fosil dinosaurus dengan cetakan kulit bersisik telah ditemukan sejak abad ke-19. Samuel Beckles menemukan tungkai depan sauropoda dengan kulit yang terawetkan pada tahun 1852 yang secara keliru dianggap berasal dari buaya; temuan tersebut dikoreksi oleh Marsh pada tahun 1888 dan menjadi subjek penelitian lebih lanjut oleh Reginald Hooley pada tahun 1917.[73] Di antara ornithischia, pada tahun 1884 Jacob Wortman menemukan cetakan kulit pada spesimen pertama Edmontosaurus annectens yang diketahui, yang sebagian besar hancur selama penggalian spesimen tersebut.[74] Owen dan Hooley kemudian mendeskripsikan cetakan kulit Hypsilophodon dan Iguanodon pada tahun 1885 dan 1917.[73] Sejak saat itu, cetakan sisik paling sering ditemukan di kalangan hadrosaurida, di mana cetakan tersebut diketahui mencakup hampir seluruh tubuh pada berbagai spesimen.[75]


Mulai tahun 1990-an, penemuan besar fosil yang terawetkan secara luar biasa dalam endapan yang dikenal sebagai Lagerstätten konservasi berkontribusi pada penelitian mengenai jaringan lunak dinosaurus.[76][77] Terutama di antaranya adalah batuan yang menghasilkan biota Jehol (Kapur Awal) dan Yanliao (Jura Tengah hingga Akhir) di timur laut Tiongkok, yang darinya ratusan spesimen dinosaurus yang memuat cetakan struktur mirip bulu (baik yang berkerabat dekat dengan burung maupun tidak, lihat § Asal-usul burung) telah dideskripsikan oleh Xu Xing dan rekan-rekannya.[78][79] Pada reptil dan mamalia yang hidup, struktur seluler penyimpan pigmen yang dikenal sebagai melanosom sebagian bertanggung jawab untuk menghasilkan pewarnaan.[80][81] Baik jejak kimia melanin maupun melanosom dengan bentuk yang khas telah dilaporkan dari bulu dan sisik dinosaurus Jehol dan Yanliao, termasuk theropoda dan ornithischia.[82] Hal ini memungkinkan berbagai rekonstruksi tubuh lengkap dari pewarnaan dinosaurus, seperti untuk Sinosauropteryx[83] dan Psittacosaurus[84] oleh Jakob Vinther dan rekan-rekannya, dan teknik serupa juga telah diperluas ke fosil dinosaurus dari lokasi lain.[80] (Namun, beberapa peneliti juga menyarankan bahwa melanosom yang terfosilisasi mewakili sisa-sisa bakteri.[85][86]) Isi perut pada beberapa dinosaurus Jehol dan Yanliao yang berkerabat dekat dengan burung juga memberikan indikasi tidak langsung mengenai pola makan dan anatomi sistem pencernaan (misalnya, tembolok).[87][88] Bukti anatomi internal yang lebih konkret telah dilaporkan pada Scipionyx dari Pietraroja Plattenkalk di Italia. Fosil ini mengawetkan bagian usus, kolon, hati, otot, dan tenggorokan.[89]

Secara bersamaan, serangkaian pekerjaan yang dipimpin oleh Mary Higby Schweitzer, Jack Horner, dan rekan-rekannya melaporkan berbagai kejadian jaringan lunak dan protein yang terawetkan di dalam fosil tulang dinosaurus. Berbagai struktur terimineralisasi yang kemungkinan mewakili sel darah merah dan serat kolagen telah ditemukan oleh Schweitzer dan lainnya dalam tulang tyrannosauridae sejak tahun 1991.[90][91][92] Namun, pada tahun 2005, Schweitzer dan rekan-rekannya melaporkan bahwa femur Tyrannosaurus mengawetkan jaringan lunak dan fleksibel di dalamnya, termasuk pembuluh darah, matriks tulang, dan jaringan ikat (serat tulang) yang masih mempertahankan struktur mikroskopisnya.[93]
Penemuan ini menunjukkan bahwa jaringan lunak asli dapat terawetkan selama waktu geologis,[72] dengan berbagai mekanisme yang telah diusulkan.[94] Kemudian, pada tahun 2009, Schweitzer dan rekan-rekannya melaporkan bahwa femur Brachylophosaurus mengawetkan struktur mikro serupa, dan teknik imunohistokimia (berdasarkan pengikatan antibodi) menunjukkan keberadaan protein seperti kolagen, elastin, dan laminin.[95] Kedua spesimen menghasilkan urutan protein kolagen yang layak untuk analisis filogenetik molekuler, yang mengelompokkan mereka dengan burung sebagaimana diperkirakan.[95][96] Ekstraksi DNA yang terfragmentasi juga telah dilaporkan untuk kedua fosil ini,[97] bersama dengan spesimen Hypacrosaurus.[98] Pada tahun 2015, Sergio Bertazzo dan rekan-rekannya melaporkan pengawetan serat kolagen dan sel darah merah pada delapan spesimen dinosaurus Kapur yang tidak menunjukkan tanda-tanda pengawetan luar biasa, yang mengindikasikan bahwa jaringan lunak mungkin terawetkan lebih umum daripada yang diperkirakan sebelumnya.[99] Dugaan bahwa struktur ini mewakili biofilm bakteri[100] telah ditolak,[101] namun kontaminasi silang tetap menjadi kemungkinan yang sulit dideteksi.[102]
Sejarah evolusi
suntingAsal-usul dan evolusi awal
sunting
Dinosaurus memisahkan diri dari leluhur archosauria mereka selama kala Trias Tengah hingga Akhir, sekitar 20 juta tahun setelah peristiwa kepunahan Perm–Trias yang dahsyat memusnahkan sekitar 96% dari semua spesies laut dan 70% spesies vertebrata darat sekitar 252 juta tahun lalu.[103][104] Fosil dinosaurus tertua yang diketahui dari sisa-sisa yang substansial berasal dari kala Karnian pada periode Trias dan ditemukan terutama di Ischigualasto dan Formasi Santa Maria di Argentina dan Brasil, serta Formasi Pebbly Arkose di Zimbabwe.[105]
Formasi Ischigualasto (yang bertanggal secara radiometrik berusia 231–230 juta tahun[106]) telah menghasilkan saurischia awal Eoraptor, yang awalnya dianggap sebagai anggota Herrerasauridae[107] tetapi kini dianggap sebagai sauropodomorpha awal, bersama dengan herrerasauridae Herrerasaurus dan Sanjuansaurus, serta sauropodomorpha Chromogisaurus, Eodromaeus, dan Panphagia.[108] Kemungkinan kemiripan Eoraptor dengan leluhur bersama semua dinosaurus menunjukkan bahwa dinosaurus pertama adalah predator bipedal berukuran kecil.[109][110][111] Formasi Santa Maria (yang bertanggal secara radiometrik lebih tua, pada 233,23 juta tahun[112]) telah menghasilkan herrerasauridae Gnathovorax dan Staurikosaurus, bersama dengan sauropodomorpha Bagualosaurus, Buriolestes, Guaibasaurus, Macrocollum, Nhandumirim, Pampadromaeus, Saturnalia, dan Unaysaurus.[108] Formasi Pebbly Arkose, yang usianya tidak pasti tetapi kemungkinan sebanding dengan dua formasi lainnya, telah menghasilkan sauropodomorpha Mbiresaurus, bersama dengan herrerasauridae yang belum diberi nama.[105]
Sisa-sisa yang kurang terawetkan dengan baik dari sauropodomorpha Jaklapallisaurus dan Nambalia, serta saurischia awal Alwalkeria, diketahui dari Formasi Maleri Atas dan Formasi Maleri Bawah di India.[113] Formasi Chañares di Argentina yang berusia Karnian mengawetkan ornithodira primitif mirip dinosaurus seperti Lagosuchus dan Lagerpeton di Argentina, menjadikannya situs penting lain untuk memahami evolusi dinosaurus. Ornithodira ini mendukung model dinosaurus awal sebagai predator bipedal berukuran kecil.[108][114] Dinosaurus mungkin telah muncul sedini kala Anisian dari periode Trias, sekitar 243 juta tahun yang lalu, yang merupakan usia Nyasasaurus dari Formasi Manda di Tanzania. Namun, fosil yang diketahui terlalu terfragmentasi untuk mengidentifikasinya sebagai dinosaurus atau hanya kerabat dekat.[115] Penunjukan Formasi Manda ke kala Anisian juga tidak pasti. Terlepas dari itu, dinosaurus hidup berdampingan dengan ornithodira non-dinosaurus selama periode waktu tertentu, dengan perkiraan berkisar antara 5–10 juta tahun[116] hingga 21 juta tahun.[112]
Ketika dinosaurus muncul, mereka bukanlah hewan darat yang dominan. Habitat terestrial ditempati oleh berbagai jenis Archosauromorpha dan therapsida, seperti cynodontia dan rhynchosauria. Pesaing utama mereka adalah pseudosuchia, seperti aetosauria, ornithosuchidae, dan rauisuchia, yang lebih sukses daripada dinosaurus.[117] Sebagian besar hewan lain ini punah pada periode Trias, dalam salah satu dari dua peristiwa. Pertama, sekitar 215 juta tahun lalu, berbagai archosauromorpha basal, termasuk protosauria, menjadi punah. Ini diikuti oleh peristiwa kepunahan Trias–Jura (sekitar 201 juta tahun lalu), yang mengakhiri sebagian besar kelompok archosauria awal lainnya, seperti aetosauria, ornithosuchidae, phytosauria, dan rauisuchia. Rhynchosauria dan dicynodontia bertahan (setidaknya di beberapa daerah) paling tidak hingga awal–pertengahan Norian dan akhir Norian atau tingkat Rhaetian paling awal,[118][119] dan tanggal pasti kepunahan mereka tidak pasti. Kehilangan ini meninggalkan fauna darat yang terdiri dari crocodylomorpha, dinosaurus, mamalia, pterosauria, dan kura-kura.[7] Beberapa garis keturunan pertama dinosaurus berdiversifikasi melalui tingkat Karnian dan Norian dari periode Trias, kemungkinan dengan menempati relung dari kelompok-kelompok yang telah punah.[9] Juga yang patut dicatat, terdapat peningkatan laju kepunahan selama episode pluvial Karnian.[120]
Evolusi dan paleobiogeografi
sunting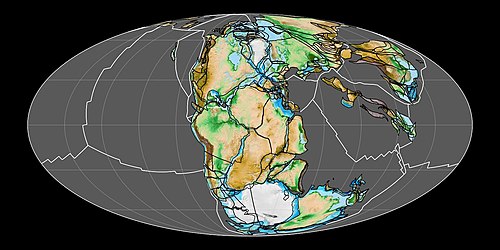
Evolusi dinosaurus setelah Trias mengikuti perubahan vegetasi dan lokasi benua. Pada Trias Akhir dan Jura Awal, benua-benua terhubung sebagai satu daratan tunggal Pangaea, dan terdapat fauna dinosaurus di seluruh dunia yang sebagian besar terdiri dari karnivora coelophysoidea dan herbivora sauropodomorpha awal.[121] Tumbuhan Gymnospermae (khususnya runjung), yang merupakan sumber makanan potensial, beradiasi pada Trias Akhir. Sauropodomorpha awal tidak memiliki mekanisme canggih untuk memproses makanan di dalam mulut, sehingga harus menggunakan cara lain untuk memecah makanan lebih jauh di sepanjang saluran pencernaan.[122] Homogenitas umum fauna dinosaurus berlanjut hingga Jura Tengah dan Akhir, di mana sebagian besar lokasi memiliki predator yang terdiri dari ceratosauria, megalosauroida, dan allosauroida, serta herbivora yang terdiri dari ornithischia stegosauria dan sauropoda besar. Contohnya termasuk Formasi Morrison di Amerika Utara dan Lapisan Tendaguru di Tanzania. Dinosaurus di Tiongkok menunjukkan beberapa perbedaan, dengan theropoda metriacanthosauridae yang terspesialisasi dan sauropoda berleher panjang yang tidak biasa seperti Mamenchisaurus.[121] Ankylosauria dan ornithopoda juga menjadi lebih umum, tetapi sauropodomorpha primitif telah punah. Tumbuhan runjung dan pteridophyta adalah tanaman yang paling umum. Sauropoda, seperti sauropodomorpha sebelumnya, bukanlah pemroses makanan oral, tetapi ornithischia berevolusi dengan berbagai cara untuk menangani makanan di dalam mulut, termasuk organ potensial menyerupai pipi untuk menahan makanan di dalam mulut, dan gerakan rahang untuk menggiling makanan.[122] Peristiwa evolusi penting lainnya pada periode Jura adalah munculnya burung sejati, yang merupakan keturunan dari coelurosauria maniraptora.[11]
Menjelang Kapur Awal dan berlanjutnya pemecahan Pangaea , dinosaurus menjadi sangat terdeferensiasi berdasarkan daratan. Bagian paling awal dari masa ini menyaksikan penyebaran ankylosauria, iguanodontia, dan brachiosauridae melalui Eropa, Amerika Utara, dan Afrika bagian utara. Kelompok-kelompok ini kemudian dilengkapi atau digantikan di Afrika oleh theropoda spinosauridae dan carcharodontosauridae yang besar, serta sauropoda rebbachisauridae dan titanosauria, yang juga ditemukan di Amerika Selatan. Di Asia, coelurosauria maniraptora seperti dromaeosauridae, troodontidae, dan oviraptorosauria menjadi theropoda yang umum, serta ankylosauridae dan ceratopsia awal seperti Psittacosaurus menjadi herbivora penting. Sementara itu, Australia menjadi rumah bagi fauna ankylosauria basal, hypsilophodont, dan iguanodontia.[121] Stegosauria tampaknya telah punah pada suatu titik di akhir Kapur Awal atau awal Kapur Akhir. Perubahan besar pada Kapur Awal, yang akan semakin menguat pada Kapur Akhir, adalah evolusi tumbuhan berbunga. Pada saat yang sama, beberapa kelompok herbivora dinosaurus mengembangkan cara yang lebih canggih untuk memproses makanan secara oral. Ceratopsia mengembangkan metode pengirisan dengan gigi yang bertumpuk satu sama lain dalam baterai, dan iguanodontia menyempurnakan metode penggilingan dengan baterai gigi , yang mencapai puncaknya pada hadrosauridae.[122] Beberapa sauropoda juga mengembangkan baterai gigi, yang dicontohkan dengan baik oleh rebbachisauridae Nigersaurus.[123]
Terdapat tiga fauna dinosaurus umum pada Kapur Akhir. Di benua utara Amerika Utara dan Asia, theropoda utamanya adalah tyrannosauridae dan berbagai jenis theropoda maniraptora yang lebih kecil, dengan himpunan herbivora yang didominasi ornithischia seperti hadrosauridae, ceratopsia, ankylosauridae, dan pachycephalosauria. Di benua selatan yang membentuk superbenua Gondwana yang kini terpecah, abelisauridae adalah theropoda yang umum, dan sauropoda titanosauria adalah herbivora yang umum. Terakhir, di Eropa, dromaeosauridae, iguanodontia rhabdodontidae, ankylosauria nodosauridae, dan sauropoda titanosauria lazim ditemukan.[121] Tumbuhan berbunga mengalami radiasi besar-besaran,[122] dengan rerumputan pertama muncul pada akhir Kapur.[124] Hadrosauridae penggiling dan ceratopsia pengiris menjadi sangat beragam di seluruh Amerika Utara dan Asia. Theropoda juga beradiasi sebagai herbivora atau omnivora, dengan therizinosauria dan ornithomimosauria menjadi umum.[122]
Peristiwa kepunahan Kapur–Paleogen, yang terjadi sekitar 66 juta tahun lalu pada akhir Kapur, menyebabkan kepunahan semua kelompok dinosaurus kecuali burung neornithes. Beberapa kelompok diapsida lain, termasuk crocodilia, dyrosauridae, sebecosuchia, kura-kura, kadal, ular, sphenodontia, dan choristodera, juga selamat dari peristiwa tersebut.[125]
Garis keturunan burung neornithes yang bertahan, termasuk leluhur ratite modern, bebek dan ayam, serta berbagai burung air, berdiversifikasi dengan cepat pada awal periode Paleogen, memasuki relung ekologi yang ditinggalkan kosong oleh kepunahan kelompok dinosaurus Mesozoikum seperti enantiornithes arboreal, hesperornithes akuatik, dan bahkan theropoda darat yang lebih besar (dalam bentuk Gastornis, eogruiidae, bathornithidae, ratite, geranoididae, mihirung, dan "burung teror"). Sering dinyatakan bahwa mamalia mengalahkan neornithes dalam persaingan untuk mendominasi sebagian besar relung terestrial, tetapi banyak dari kelompok ini hidup berdampingan dengan fauna mamalia yang kaya selama sebagian besar Era Kenozoikum.[126] Burung teror dan bathornithidae menempati gilda karnivora bersama mamalia predator,[127][128] dan ratite masih cukup sukses sebagai herbivora berukuran sedang; eogruiidae juga bertahan dari Eosen hingga Pliosen, dan baru punah sangat baru-baru ini setelah lebih dari 20 juta tahun hidup berdampingan dengan banyak kelompok mamalia.[129]
Klasifikasi
sunting


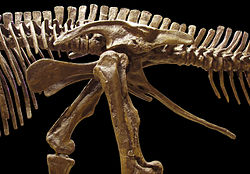
Dinosaurus termasuk dalam kelompok yang dikenal sebagai archosauria, yang juga mencakup buaya modern. Dalam kelompok archosauria, dinosaurus dibedakan paling nyata dari gaya berjalannya. Kaki dinosaurus menjulur langsung di bawah tubuh, sedangkan kaki kadal dan buaya merentang ke samping.[26]
Secara kolektif, dinosaurus sebagai klad dibagi menjadi dua cabang utama, Saurischia dan Ornithischia. Saurischia mencakup taksa yang berbagi leluhur bersama yang lebih baru dengan burung daripada dengan Ornithischia, sementara Ornithischia mencakup semua taksa yang berbagi leluhur bersama yang lebih baru dengan Triceratops daripada dengan Saurischia. Secara anatomis, kedua kelompok ini dapat dibedakan paling jelas dari struktur panggul mereka. Saurischia awal—"berpanggul kadal", dari bahasa Yunani sauros (σαῦρος) yang berarti "kadal" dan ischion (ἰσχίον) yang berarti "sendi panggul"—mempertahankan struktur panggul leluhur mereka, dengan tulang pubis yang mengarah ke kranial, atau depan.[33] Bentuk dasar ini dimodifikasi dengan rotasi pubis ke belakang dalam berbagai tingkatan pada beberapa kelompok (Herrerasaurus,[130] therizinosauroida,[131] dromaeosauridae,[132] dan burung[11]). Saurischia mencakup theropoda (secara eksklusif bipedal dan dengan berbagai jenis makanan) dan sauropodomorpha (herbivora berleher panjang yang mencakup kelompok kuadrupedal yang maju).[25][133]
Sebaliknya, ornithischia—"berpanggul burung", dari bahasa Yunani ornis (ὀρνίς) yang berarti "burung" dan ischion (ἰσχίον) yang berarti "sendi panggul"—memiliki panggul yang secara superfisial menyerupai panggul burung: tulang pubis berorientasi ke kaudal (mengarah ke belakang). Tidak seperti burung, pubis ornithischia biasanya juga memiliki prosesus (unjuran) tambahan yang mengarah ke depan. Ornithischia mencakup berbagai spesies yang sebagian besar merupakan herbivora.
Meskipun menggunakan istilah "panggul burung" (Ornithischia) dan "panggul kadal" (Saurischia), burung bukan bagian dari Ornithischia. Burung justru termasuk dalam Saurischia, dinosaurus "berpanggul kadal"—burung berevolusi dari dinosaurus awal dengan "panggul kadal".[26]
Kladogram berikut menunjukkan hubungan kelompok dinosaurus yang disederhanakan menurut Michael Benton, 2015:[33]
| Dinosauria | |
Paleobiologi
suntingPengetahuan mengenai dinosaurus diperoleh dari beragam rekam fosil dan non-fosil, meliputi tulang-belulang yang terfosilisasi, feses, jalur jejak, gastrolit, bulu, cetakan kulit, organ dalam, serta jaringan lunak lainnya.[89][93] Banyak bidang studi berkontribusi pada pemahaman kita tentang dinosaurus, termasuk fisika (terutama biomekanika), kimia, biologi, dan ilmu kebumian (di mana paleontologi adalah salah satu sub-disiplinnya).[134][135] Dua topik yang menjadi minat dan kajian khusus adalah ukuran dan perilaku dinosaurus.[136]
Ukuran
sunting
Bukti terkini menunjukkan bahwa ukuran rata-rata dinosaurus bervariasi sepanjang Trias, Jura Awal, Jura Akhir, dan Kapur.[110] Dinosaurus theropoda predator, yang menempati sebagian besar relung karnivora terestrial selama Mesozoikum, paling sering masuk dalam kategori 100-hingga-1.000 kg (220-hingga-2.200 pon) ketika diurutkan berdasarkan perkiraan berat ke dalam kategori berdasarkan orde besaran, sedangkan mamalia karnivora predator terkini memuncak pada kategori 10-hingga-100 kg (22-hingga-220 pon).[137] Modus massa tubuh dinosaurus Mesozoikum berada di antara 1 dan 10 ton metrik (1,1 dan 11,0 ton pendek).[138] Hal ini sangat kontras dengan ukuran rata-rata mamalia Kenozoikum, yang diperkirakan oleh Museum Sejarah Alam Nasional sekitar 2 hingga 5 kg (4,4 hingga 11,0 pon).[139]
Sauropoda adalah dinosaurus terbesar dan terberat. Untuk sebagian besar era dinosaurus, sauropoda terkecil lebih besar dari apa pun di habitat mereka, dan yang terbesar memiliki orde besaran yang lebih masif daripada apa pun yang pernah berjalan di Bumi sejak saat itu. Mamalia prasejarah raksasa seperti Paraceratherium (mamalia darat terbesar yang pernah ada) tampak kerdil dibandingkan dengan sauropoda raksasa, dan hanya paus modern yang mendekati atau melampaui ukuran mereka.[140] Terdapat beberapa keuntungan yang diusulkan bagi ukuran besar sauropoda, termasuk perlindungan dari predasi, pengurangan penggunaan energi, dan umur panjang, tetapi mungkin keuntungan yang paling penting adalah terkait pola makan. Hewan besar lebih efisien dalam pencernaan dibandingkan hewan kecil, karena makanan menghabiskan lebih banyak waktu dalam sistem pencernaan mereka. Hal ini juga memungkinkan mereka untuk hidup dari makanan dengan nilai gizi lebih rendah daripada hewan yang lebih kecil. Sisa-sisa sauropoda sebagian besar ditemukan dalam formasi batuan yang ditafsirkan sebagai lingkungan kering atau kering secara musiman, dan kemampuan untuk memakan dedaunan rendah nutrisi dalam jumlah besar akan menguntungkan di lingkungan seperti itu.[141]
Terbesar dan terkecil
suntingPara ilmuwan mungkin tidak akan pernah yakin mengenai dinosaurus terbesar dan terkecil yang pernah ada. Hal ini karena hanya sebagian kecil hewan yang terfosilisasi dan sebagian besar dari mereka tetap terkubur di dalam bumi. Sedikit spesimen dinosaurus non-avian yang ditemukan berupa kerangka lengkap, dan cetakan kulit serta jaringan lunak lainnya jarang ditemukan. Membangun kembali kerangka lengkap dengan membandingkan ukuran dan morfologi tulang dengan spesies serupa yang lebih dikenal adalah seni yang tidak pasti, dan merekonstruksi otot serta organ lain dari hewan hidup adalah, paling banter, sebuah proses penebakan yang terpelajar.[142]

Dinosaurus tertinggi dan terberat yang diketahui dari kerangka yang baik adalah Giraffatitan brancai (sebelumnya diklasifikasikan sebagai spesies Brachiosaurus). Sisa-sisanya ditemukan di Tanzania antara tahun 1907 dan 1912. Tulang-tulang dari beberapa individu berukuran serupa digabungkan ke dalam kerangka yang sekarang dipasang dan dipajang di Museum für Naturkunde di Berlin;[143] kerangka pajangan ini tingginya 12 meter (39 ft) dan panjangnya 218 hingga 225 meter (715 hingga 738 ft),[144][145] dan mungkin milik hewan yang beratnya antara 30000 dan 60000 kilogram (70000 dan 130000 lb). Dinosaurus lengkap terpanjang adalah Diplodocus sepanjang 27 meter (89 ft), yang ditemukan di Wyoming di Amerika Serikat dan dipajang di Museum Sejarah Alam Carnegie di Pittsburgh pada tahun 1907.[146] Dinosaurus terpanjang yang diketahui dari material fosil yang baik adalah Patagotitan: kerangka pajangan di Museum Sejarah Alam Amerika di New York panjangnya 37 meter ([convert: unit tak dikenal]). Museo Municipal Carmen Funes di Plaza Huincul, Argentina, memiliki kerangka rekonstruksi Argentinosaurus yang panjangnya 397 meter ([convert: unit tak dikenal]).[147]

Ada dinosaurus yang lebih besar, tetapi pengetahuan tentang mereka sepenuhnya didasarkan pada sejumlah kecil fosil yang terfragmentasi. Sebagian besar spesimen herbivora terbesar yang tercatat ditemukan pada tahun 1970-an atau setelahnya, dan termasuk Argentinosaurus yang masif, yang mungkin berberat 80000 hingga 100000 kilogram (88 hingga 110 ton pendek) dan mencapai panjang 30 hingga 40 meter (98 hingga 131 ft); beberapa yang terpanjang adalah Diplodocus hallorum[141] (sebelumnya Seismosaurus) sepanjang 335-meter (1.099 ft), Supersaurus sepanjang 33-hingga-34-meter (108 hingga 112 ft),[148] dan Patagotitan sepanjang 37-meter ([convert: unit tak dikenal]); serta yang tertinggi, Sauroposeidon setinggi 18-meter (59 ft), yang bisa mencapai jendela lantai enam. Ada beberapa dinosaurus yang dianggap terberat atau terpanjang. Yang paling terkenal termasuk Amphicoelias fragillimus, yang hanya diketahui dari lengkung saraf vertebra parsial yang sekarang hilang yang dideskripsikan pada tahun 1878. Berdasarkan ekstrapolasi dari ilustrasi tulang ini, hewan tersebut mungkin memiliki panjang 58 meter (190 ft) dan berat 122400 kg (269800 pon).[141] Namun, penelitian terbaru telah mengklasifikasikan ulang Amphicoelias dari diplodocid yang panjang dan ramping menjadi rebbachisaurid yang lebih pendek namun jauh lebih kekar. Sekarang diganti namanya menjadi Maraapunisaurus, sauropoda ini berukuran panjang hingga 40 meter (130 ft) dan berat hingga 120000 kg (260000 pon).[149][150] Pesaing lain untuk gelar ini termasuk Bruhathkayosaurus, takson kontroversial yang baru-baru ini dikonfirmasi keberadaannya setelah foto-foto yang diarsipkan ditemukan.[151] Bruhathkayosaurus adalah titanosaurus dan kemungkinan besar berbobot lebih berat daripada Maraapunisaurus. Estimasi ukuran terbaru pada tahun 2023 menunjukkan sauropoda ini mencapai panjang hingga 44 m (144 ft) dan ditempatkan dalam kisaran berat kolosal sekitar 110000–170000 kg (240000–370000 pon). Jika perkiraan batas atas ini benar, Bruhathkayosaurus akan menyaingi paus biru dan Perucetus colossus sebagai salah satu hewan terbesar yang pernah ada.[152]
Dinosaurus karnivora terbesar adalah Spinosaurus, yang mencapai panjang 12,6 hingga 18 meter (41 hingga 59 ft) dan berat 7 hingga 20,9 ton metrik (7,7 hingga 23,0 ton pendek).[153][154] Theropoda karnivora besar lainnya termasuk Giganotosaurus, Carcharodontosaurus, dan Tyrannosaurus.[154] Therizinosaurus dan Deinocheirus adalah salah satu theropoda tertinggi. Dinosaurus ornithischia terbesar mungkin adalah hadrosauridae Shantungosaurus giganteus yang berukuran 166 meter ([convert: unit tak dikenal]).[155] Individu terbesar mungkin berbobot hingga 16 ton metrik (18 ton pendek).[156]

Dinosaurus terkecil yang diketahui adalah kolibri lebah,[157] dengan panjang hanya 5 sentimeter (2,0 in) dan massa sekitar 18 g (0,63 oz).[158] Dinosaurus non-avialan terkecil yang diketahui berukuran sekitar merpati dan merupakan theropoda yang paling dekat kekerabatannya dengan burung.[159] Sebagai contoh, Anchiornis huxleyi saat ini adalah dinosaurus non-avialan terkecil yang dideskripsikan dari spesimen dewasa, dengan perkiraan berat 110 g (3,9 oz)[160] dan total panjang kerangka 34 sentimeter (1,12 ft).[159][160] Dinosaurus non-avialan herbivora terkecil termasuk Microceratus dan Wannanosaurus, masing-masing dengan panjang sekitar 60 sentimeter (2,0 ft).[161][162]
Perilaku
sunting
Banyak burung modern sangat sosial, sering ditemukan hidup dalam kawanan. Ada kesepakatan umum bahwa beberapa perilaku yang umum pada burung, serta pada crocodilia (kerabat hidup terdekat burung), juga umum di antara kelompok dinosaurus yang telah punah. Interpretasi perilaku pada spesies fosil umumnya didasarkan pada pose kerangka dan habitatnya, simulasi komputer dari biomekanika mereka, dan perbandingan dengan hewan modern di relung ekologi yang serupa.[134]
Bukti potensial pertama untuk perilaku menggembala atau berkelompok sebagai perilaku luas yang umum bagi banyak kelompok dinosaurus selain burung adalah penemuan 31 Iguanodon pada tahun 1878, ornithischia yang saat itu diperkirakan tewas bersama di Bernissart, Belgia, setelah mereka jatuh ke dalam lubang amblas yang dalam dan banjir lalu tenggelam.[163] Situs kematian massal lainnya telah ditemukan kemudian. Situs-situs tersebut, bersama dengan beberapa jalur jejak, menunjukkan bahwa perilaku suka berteman adalah hal yang umum pada banyak spesies dinosaurus awal. Jalur jejak ratusan atau bahkan ribuan herbivora menunjukkan bahwa dinosaurus paruh bebek (hadrosaurida) mungkin bergerak dalam kawanan besar, seperti bison Amerika atau springbok Afrika. Jejak sauropoda mendokumentasikan bahwa hewan-hewan ini bepergian dalam kelompok yang terdiri dari beberapa spesies berbeda, setidaknya di Oxfordshire, Inggris,[164] meskipun tidak ada bukti untuk struktur kawanan tertentu.[165] Berkumpul dalam kawanan mungkin telah berevolusi untuk pertahanan, untuk tujuan migrasi, atau untuk memberikan perlindungan bagi yang muda. Ada bukti bahwa banyak jenis dinosaurus yang tumbuh lambat, termasuk berbagai theropoda, sauropoda, ankylosauria, ornithopoda, dan ceratopsia, membentuk agregasi individu yang belum dewasa. Salah satu contohnya adalah situs di Mongolia Dalam yang telah menghasilkan sisa-sisa lebih dari 20 Sinornithomimus, berusia satu hingga tujuh tahun. Kumpulan ini ditafsirkan sebagai kelompok sosial yang terjebak dalam lumpur.[166] Interpretasi dinosaurus sebagai hewan yang suka berteman juga telah meluas hingga menggambarkan theropoda karnivora sebagai pemburu kawanan yang bekerja sama untuk menjatuhkan mangsa besar.[167][168] Namun, gaya hidup ini jarang terjadi di antara burung modern, buaya, dan reptil lainnya, dan bukti tafonomi yang menyarankan perburuan kawanan mirip mamalia pada theropoda seperti Deinonychus dan Allosaurus juga dapat ditafsirkan sebagai hasil dari perselisihan fatal antara hewan yang sedang makan, seperti yang terlihat pada banyak predator diapsida modern.[169]

Jambul dan jumbai dari beberapa dinosaurus, seperti marginocephalia, theropoda, dan lambeosaurinae, mungkin terlalu rapuh untuk digunakan sebagai pertahanan aktif, sehingga kemungkinan besar digunakan untuk peragaan seksual atau agresif, meskipun sedikit yang diketahui tentang perkawinan dan teritorialisme dinosaurus. Luka di kepala akibat gigitan menunjukkan bahwa theropoda, setidaknya, terlibat dalam konfrontasi agresif aktif.[170]
Dari sudut pandang perilaku, salah satu fosil dinosaurus yang paling berharga ditemukan di Gurun Gobi pada tahun 1971. Fosil tersebut mencakup seekor Velociraptor yang sedang menyerang seekor Protoceratops,[171] memberikan bukti bahwa dinosaurus memang saling menyerang.[172] Bukti tambahan untuk penyerangan mangsa hidup adalah ekor yang sembuh sebagian dari seekor Edmontosaurus, dinosaurus hadrosaurida; ekornya rusak sedemikian rupa sehingga menunjukkan bahwa hewan tersebut digigit oleh tyrannosaur namun selamat.[172] Kanibalisme di antara beberapa spesies dinosaurus dikonfirmasi oleh bekas gigi yang ditemukan di Madagaskar pada tahun 2003, yang melibatkan theropoda Majungasaurus.[173]
Perbandingan antara cincin sklerotik dinosaurus serta burung dan reptil modern telah digunakan untuk menyimpulkan pola aktivitas harian dinosaurus. Meskipun telah disarankan bahwa sebagian besar dinosaurus aktif pada siang hari, perbandingan ini menunjukkan bahwa dinosaurus predator kecil seperti dromaeosauridae, Juravenator, dan Megapnosaurus kemungkinan bersifat nokturnal. Dinosaurus herbivora dan omnivora berukuran besar dan sedang seperti ceratopsia, sauropodomorpha, hadrosaurida, ornithomimosauria mungkin bersifat katemeral, aktif selama interval singkat sepanjang hari, meskipun ornithischia kecil Agilisaurus disimpulkan bersifat diurnal.[174]
Berdasarkan bukti fosil dari dinosaurus seperti Oryctodromeus, beberapa spesies ornithischia tampaknya menjalani gaya hidup sebagian fosorial (menggali liang).[175] Banyak burung modern bersifat arboreal (memanjat pohon), dan hal ini juga berlaku bagi banyak burung Mesozoikum, terutama enantiornithes.[176] Meskipun beberapa spesies mirip burung awal mungkin juga sudah arboreal (termasuk dromaeosauridae) seperti Microraptor[177]) sebagian besar dinosaurus non-avialan tampaknya mengandalkan lokomosi berbasis darat. Pemahaman yang baik tentang bagaimana dinosaurus bergerak di tanah adalah kunci untuk model perilaku dinosaurus; ilmu biomekanika, yang dipelopori oleh Robert McNeill Alexander, telah memberikan wawasan yang signifikan di bidang ini. Misalnya, studi tentang gaya yang diberikan oleh otot dan gravitasi pada struktur kerangka dinosaurus telah menyelidiki seberapa cepat dinosaurus dapat berlari,[134] apakah diplodocidae dapat menciptakan dentuman sonik melalui sentakan ekor seperti cambuk,[178] dan apakah sauropoda dapat mengapung.[179]
Komunikasi
sunting
Burung modern berkomunikasi dengan sinyal visual dan pendengaran, dan keragaman luas struktur peragaan visual di antara kelompok dinosaurus fosil, seperti tanduk, jumbai, jambul, layar, dan bulu, menunjukkan bahwa komunikasi visual selalu penting dalam biologi dinosaurus.[180] Rekonstruksi warna bulu Anchiornis menyarankan pentingnya warna dalam komunikasi visual pada dinosaurus non-avian.[181] Sebuah tinjauan tahun 2009 mengindikasikan bahwa dinosaurus non-avian menggunakan peragaan visual dan mungkin suara non-vokal, seperti mendesis, menggertakkan atau mengatupkan rahang, memercikkan air, dan mengepakkan sayap (mungkin pada dinosaurus maniraptora bersayap).[180]
Laring atau kotak suara yang terfosilisasi telah diidentifikasi pada dua dinosaurus; ankylosauridae Pinacosaurus dan neornithischia Pulaosaurus, yang strukturnya menunjukkan bahwa dinosaurus mampu menghasilkan vokalisasi kompleks seperti burung.[182][183] Sebuah studi tahun 2016 menyimpulkan bahwa beberapa dinosaurus mungkin menghasilkan vokalisasi mulut tertutup, seperti mendekut, berseru, dan menderu. Ini terjadi pada reptil dan burung serta melibatkan pengembangan esofagus atau kantung trakea. Vokalisasi semacam itu berevolusi secara independen pada archosauria yang masih ada berkali-kali, mengikuti peningkatan ukuran tubuh.[184] Jambul beberapa hadrosaurida dan ruang hidung ankylosauridae mungkin merupakan resonator.[185][186] Burung telah mengembangkan organ baru untuk vokalisasi, siring,[187] sisa-sisa paling awal yang ditemukan dalam spesimen Vegavis iaai yang mirip bebek yang bertanggal 69 –66 juta tahun lalu.[188] Siring mungkin telah melengkapi dan kemudian menggantikan laring sebagai organ vokal selama evolusi burung.[189]
Biologi reproduksi
sunting
Semua dinosaurus menghasilkan telur amniotik. Telur dinosaurus biasanya diletakkan di dalam sarang. Sebagian besar spesies membuat sarang yang agak rumit yang dapat berupa mangkuk, kubah, piringan, lekukan tanah, gundukan, atau liang.[190] Beberapa spesies burung modern tidak memiliki sarang; guillemot biasa yang bersarang di tebing meletakkan telurnya di atas batu gundul, dan penguin kaisar jantan menyimpan telur di antara tubuh dan kaki mereka. Burung primitif dan banyak dinosaurus non-avian sering kali bertelur di sarang komunal, dengan jantan yang terutama mengerami telur-telur tersebut. Sementara burung modern hanya memiliki satu oviduk fungsional dan mengeluarkan satu telur dalam satu waktu, burung yang lebih primitif dan dinosaurus memiliki dua oviduk, seperti buaya. Beberapa dinosaurus non-avian, seperti Troodon, menunjukkan pola bertelur iteratif, di mana induk mungkin mengeluarkan sepasang telur setiap satu atau dua hari, dan kemudian memastikan penetasan serentak dengan menunda pengeraman hingga semua telur dikeluarkan.[191]
Saat bertelur, betina menumbuhkan jenis tulang khusus di antara tulang luar yang keras dan sumsum pada tungkai mereka. Tulang meduler ini, yang kaya akan kalsium, digunakan untuk membuat cangkang telur. Penemuan fitur-fitur pada kerangka Tyrannosaurus memberikan bukti adanya tulang meduler pada dinosaurus yang telah punah dan, untuk pertama kalinya, memungkinkan paleontolog untuk menentukan jenis kelamin spesimen fosil dinosaurus. Penelitian lebih lanjut menemukan tulang meduler pada carnosauria Allosaurus dan ornithopoda Tenontosaurus. Karena garis keturunan dinosaurus yang mencakup Allosaurus dan Tyrannosaurus memisahkan diri dari garis keturunan yang mengarah ke Tenontosaurus sangat awal dalam evolusi dinosaurus, hal ini menunjukkan bahwa produksi jaringan meduler adalah karakteristik umum dari semua dinosaurus.[192]

Sifat lain yang tersebar luas di antara burung modern (namun lihat di bawah sehubungan dengan kelompok fosil dan megapoda yang masih ada) adalah pengasuhan induk terhadap anak setelah menetas. Penemuan area peneluran Maiasaura ("kadal ibu yang baik") di Montana pada tahun 1978 oleh Jack Horner menunjukkan bahwa pengasuhan induk berlanjut lama setelah kelahiran di antara ornithopoda.[193] Sebuah spesimen oviraptorid Citipati osmolskae ditemukan dalam posisi mengeram mirip ayam pada tahun 1993,[194] yang mungkin mengindikasikan bahwa mereka mulai menggunakan lapisan bulu isolator untuk menjaga telur tetap hangat.[195] Sebuah embrio sauropodomorpha basal Massospondylus ditemukan tanpa gigi, yang mengindikasikan bahwa diperlukan pengasuhan induk untuk memberi makan dinosaurus muda tersebut.[196] Jalur jejak juga telah mengonfirmasi perilaku pengasuhan di antara ornithopoda dari Pulau Skye di barat laut Skotlandia.[197]
Namun, terdapat banyak bukti mengenai prekosialitas atau superprekosialitas di antara banyak spesies dinosaurus, khususnya theropoda. Sebagai contoh, burung non-ornithuromorpha telah secara melimpah terbukti memiliki laju pertumbuhan yang lambat, perilaku mengubur telur seperti megapoda, dan kemampuan terbang segera setelah lahir.[198][199][200][201] Baik Tyrannosaurus maupun Troodon memiliki juvenil dengan superprekosialitas yang jelas dan kemungkinan menempati relung ekologi yang berbeda dari dewasa.[191] Superprekosialitas telah disimpulkan juga terjadi pada sauropoda.[202]
Struktur genital tidak mungkin terfosilisasi karena tidak memiliki sisik yang memungkinkan pengawetan melalui pigmentasi atau sisa garam kalsium fosfat. Pada tahun 2021, spesimen eksterior lubang kloaka dinosaurus yang paling terawetkan dideskripsikan untuk Psittacosaurus, menunjukkan pembengkakan lateral yang mirip dengan kelenjar kesturi buaya yang digunakan dalam peragaan sosial oleh kedua jenis kelamin dan daerah berpigmen yang juga dapat mencerminkan fungsi sinyal. Namun, spesimen ini sendiri tidak menawarkan cukup informasi untuk menentukan apakah dinosaurus ini memiliki fungsi sinyal seksual; ia hanya mendukung kemungkinannya. Sinyal visual kloaka dapat terjadi pada jantan atau betina pada burung yang masih hidup, sehingga kecil kemungkinannya berguna untuk menentukan jenis kelamin dinosaurus yang telah punah.[203]
Fisiologi
suntingKarena buaya dan burung modern sama-sama memiliki jantung beruang empat (meskipun termodifikasi pada buaya), kemungkinan besar ini adalah ciri yang dimiliki oleh semua archosauria, termasuk semua dinosaurus.[204] Walaupun semua burung modern memiliki metabolisme tinggi dan bersifat endoterm ("berdarah panas"), perdebatan sengit telah berlangsung sejak tahun 1960-an mengenai seberapa jauh ke belakang dalam garis keturunan dinosaurus sifat ini bermula. Berbagai peneliti telah mendukung gagasan bahwa dinosaurus bersifat endotermik, ektoterm ("berdarah dingin"), atau berada di antara keduanya.[205] Konsensus yang muncul di kalangan peneliti adalah bahwa, meskipun garis keturunan dinosaurus yang berbeda akan memiliki metabolisme yang berbeda pula, sebagian besar dari mereka memiliki laju metabolisme yang lebih tinggi daripada reptil lain tetapi lebih rendah daripada burung dan mamalia yang masih hidup,[206] yang oleh beberapa pihak disebut sebagai mesotermik.[207] Bukti dari buaya dan kerabat mereka yang telah punah menunjukkan bahwa metabolisme tinggi seperti itu bisa saja berkembang pada archosauria paling awal, yang merupakan leluhur bersama dinosaurus dan buaya.[208][209]

Setelah dinosaurus non-avian ditemukan, para paleontolog awalnya mendalilkan bahwa mereka bersifat ektotermik. Hal ini digunakan untuk menyiratkan bahwa dinosaurus purba adalah organisme yang relatif lamban dan lesu, meskipun banyak reptil modern yang cepat dan lincah walaupun bergantung pada sumber panas eksternal untuk mengatur suhu tubuh mereka. Gagasan dinosaurus sebagai ektoterm tetap menjadi pandangan umum hingga Robert T. Bakker, pendukung awal endotermi dinosaurus, menerbitkan makalah berpengaruh tentang topik tersebut pada tahun 1968. Bakker secara khusus menggunakan bukti anatomi dan ekologi untuk berargumen bahwa sauropoda, yang sampai saat itu digambarkan sebagai hewan akuatik yang merayap dengan ekor menyeret di tanah, adalah endoterm yang menjalani kehidupan terestrial yang penuh semangat. Pada tahun 1972, Bakker memperluas argumennya berdasarkan kebutuhan energi dan rasio predator-mangsa. Ini adalah salah satu hasil penting yang mengarah pada renaisans dinosaurus.[59][62][63][210]
Salah satu kontribusi terbesar bagi pemahaman modern tentang fisiologi dinosaurus adalah paleohistologi, studi tentang struktur jaringan mikroskopis pada dinosaurus.[211][212] Sejak tahun 1960-an dan seterusnya, Armand de Ricqlès menyarankan bahwa keberadaan tulang fibrolamelar—jaringan tulang dengan tekstur berserat yang tidak teratur dan dipenuhi pembuluh darah—mengindikasikan pertumbuhan yang konsisten cepat dan oleh karena itu endotermi. Tulang fibrolamelar umum ditemukan baik pada dinosaurus maupun pterosaurus,[213][214] meskipun tidak selalu ada.[215][216] Hal ini telah memicu sejumlah besar pekerjaan dalam merekonstruksi kurva pertumbuhan dan memodelkan evolusi laju pertumbuhan di berbagai garis keturunan dinosaurus,[217] yang secara keseluruhan menunjukkan bahwa dinosaurus tumbuh lebih cepat daripada reptil yang masih hidup.[212] Bukti lain yang menyarankan endotermi meliputi keberadaan bulu dan jenis penutup tubuh lainnya pada banyak garis keturunan (lihat § Bulu); rasio isotop oksigen-18 yang lebih konsisten dalam jaringan tulang dibandingkan dengan ektoterm, terutama karena variasi lintang dan suhu udara, yang menunjukkan suhu internal yang stabil[218][219] (meskipun rasio ini dapat berubah selama fosilisasi[220]); serta penemuan dinosaurus kutub, yang hidup di Australia, Antartika, dan Alaska ketika tempat-tempat tersebut memiliki iklim sejuk dan sedang.[221][222][223][224]

Pada dinosaurus saurischia, metabolisme yang lebih tinggi didukung oleh evolusi sistem pernapasan burung, yang ditandai dengan sistem kantung udara yang luas yang memperluas paru-paru dan menembus banyak tulang dalam kerangka, menjadikannya berongga.[225] Sistem pernapasan seperti itu, yang mungkin muncul pada saurischia paling awal,[226] akan memberi mereka lebih banyak oksigen dibandingkan dengan mamalia berukuran sama, sembari juga memiliki volume tidal istirahat yang lebih besar dan membutuhkan frekuensi pernapasan yang lebih rendah, yang akan memungkinkan mereka untuk mempertahankan tingkat aktivitas yang lebih tinggi.[140] Aliran udara yang cepat juga akan menjadi mekanisme pendinginan yang efektif, yang dalam hubungannya dengan laju metabolisme yang lebih rendah[227] akan mencegah sauropoda besar dari kepanasan. Ciri-ciri ini mungkin telah memungkinkan sauropoda tumbuh dengan cepat hingga mencapai ukuran raksasa.[228][229] Sauropoda juga mungkin mendapat manfaat dari ukurannya—rasio luas permukaan terhadap volume yang kecil berarti mereka akan mampu melakukan termoregulasi dengan lebih mudah, sebuah fenomena yang disebut gigantotermi.[140][230]
Seperti reptil lainnya, dinosaurus pada dasarnya bersifat urikotelik, yaitu ginjal mereka mengekstraksi limbah nitrogen dari aliran darah dan mengeluarkannya sebagai asam urat, bukan urea atau amonia, melalui ureter ke dalam usus. Hal ini akan membantu mereka menghemat air.[206] Pada sebagian besar spesies yang hidup, asam urat dikeluarkan bersama dengan feses sebagai limbah semipadat.[231][232] Namun, setidaknya beberapa burung modern (seperti kolibri) dapat secara fakultatif bersifat amonotelik, mengeluarkan sebagian besar limbah nitrogen sebagai amonia.[233] Bahan ini, serta keluaran dari usus, muncul dari kloaka.[234][235] Selain itu, banyak spesies memuntahkan pelet,[236] dan pelet fosil diketahui sudah ada sejak zaman Jura dari Anchiornis.[237]
Ukuran dan bentuk otak dapat direkonstruksi sebagian berdasarkan tulang di sekitarnya. Pada tahun 1896, Marsh menghitung rasio antara berat otak dan berat badan tujuh spesies dinosaurus, yang menunjukkan bahwa otak dinosaurus secara proporsional lebih kecil daripada buaya saat ini, dan bahwa otak Stegosaurus lebih kecil daripada vertebrata darat mana pun yang masih hidup. Hal ini berkontribusi pada pandangan publik yang meluas bahwa dinosaurus itu lamban dan luar biasa bodoh. Harry Jerison, pada tahun 1973, menunjukkan bahwa otak yang secara proporsional lebih kecil memang diharapkan pada ukuran tubuh yang lebih besar, dan bahwa ukuran otak dinosaurus tidak lebih kecil dari yang diharapkan jika dibandingkan dengan reptil yang masih hidup.[238] Penelitian selanjutnya menunjukkan bahwa ukuran otak relatif meningkat secara progresif selama evolusi theropoda, dengan kecerdasan tertinggi – sebanding dengan burung modern – dihitung untuk troodontid Troodon.[239]
Asal-usul burung
suntingGagasan bahwa dinosaurus merupakan leluhur burung pertama kali dikemukakan pada tahun 1868 oleh Thomas Henry Huxley.[240] Setelah karya Gerhard Heilmann pada awal abad ke-20, teori burung sebagai keturunan dinosaurus ditinggalkan demi gagasan bahwa mereka adalah keturunan thecodontia yang digeneralisasi, dengan bukti kunci berupa dugaan ketiadaan klavikula pada dinosaurus.[241] Namun, seperti yang ditunjukkan oleh penemuan-penemuan selanjutnya, klavikula (atau tulang garpu tunggal yang menyatu, yang berasal dari klavikula yang terpisah) sebenarnya tidak absen;[11] tulang-tulang tersebut telah ditemukan sejak tahun 1924 pada Oviraptor, tetapi salah diidentifikasi sebagai interklavikula.[242] Pada tahun 1970-an, John Ostrom menghidupkan kembali teori dinosaurus–burung,[243] yang mendapatkan momentum pada dekade-dekade berikutnya dengan munculnya analisis kladistik,[244] dan peningkatan besar dalam penemuan theropoda kecil dan burung purba.[28] Catatan khusus tertuju pada fosil-fosil dari Biota Jehol, tempat berbagai theropoda dan burung purba ditemukan, sering kali dengan sejenis bulu.[11][69] Burung berbagi lebih dari seratus fitur anatomi yang berbeda dengan dinosaurus theropoda, yang sekarang secara umum diterima sebagai kerabat purba terdekat mereka.[245] Mereka berkerabat paling dekat dengan coelurosauria maniraptora.[11] Sebagian kecil ilmuwan, terutama Alan Feduccia dan Larry Martin, telah mengajukan jalur evolusi lain, termasuk versi revisi dari usulan archosauria basal Heilmann,[246] atau bahwa theropoda maniraptora adalah leluhur burung tetapi mereka sendiri bukan dinosaurus, melainkan hanya hasil evolusi konvergen dengan dinosaurus.[247]
Bulu
sunting
Bulu adalah salah satu karakteristik yang paling mudah dikenali dari burung modern, dan ciri yang juga dimiliki oleh beberapa dinosaurus non-avian. Berdasarkan distribusi bukti fosil saat ini, tampaknya bulu adalah ciri dinosaurus leluhur, meskipun ciri tersebut mungkin telah hilang secara selektif pada beberapa spesies.[248] Bukti fosil langsung berupa bulu atau struktur mirip bulu telah ditemukan pada beragam spesies di banyak kelompok dinosaurus non-avian,[69] baik di kalangan saurischia maupun ornithischia. Struktur sederhana, bercabang, dan mirip bulu diketahui dari heterodontosauridae, neornithischia primitif,[249] dan theropoda,[250] serta ceratopsia primitif. Bukti adanya bulu sejati dengan bendera yang mirip dengan bulu terbang burung modern hanya ditemukan pada subkelompok theropoda Maniraptora, yang mencakup oviraptorosauria, troodontidae, dromaeosauridae, dan burung.[11][251] Struktur mirip bulu yang dikenal sebagai piknofiber juga telah ditemukan pada pterosaurus.[252]
Namun, para peneliti tidak sepakat mengenai apakah struktur-struktur ini memiliki asal-usul yang sama antar garis keturunan (yakni, bersifat homolog),[253][254] atau apakah itu merupakan hasil dari eksperimen luas dengan penutup kulit di kalangan ornithodira.[255] Jika kasus pertama yang benar, filamen mungkin sudah umum pada garis keturunan ornithodira dan berevolusi sebelum munculnya dinosaurus itu sendiri.[248] Penelitian terhadap genetika aligator Amerika telah mengungkapkan bahwa sisik crocodilia memang memiliki keratin bulu selama perkembangan embrio, tetapi keratin ini tidak diekspresikan oleh hewan tersebut sebelum menetas.[256] Deskripsi dinosaurus berbulu secara umum tidak lepas dari kontroversi; mungkin kritikus yang paling vokal adalah Alan Feduccia dan Theagarten Lingham-Soliar, yang mengusulkan bahwa beberapa fosil yang diklaim mirip bulu adalah hasil penguraian serat kolagen yang melapisi kulit dinosaurus,[257][258][259] dan bahwa dinosaurus maniraptora dengan bulu berbendera sebenarnya bukan dinosaurus, melainkan konvergen dengan dinosaurus.[247][258] Namun, pandangan mereka sebagian besar tidak diterima oleh peneliti lain, hingga pada titik di mana sifat ilmiah dari usulan Feduccia dipertanyakan.[260]
Archaeopteryx adalah fosil pertama yang ditemukan yang mengungkapkan potensi hubungan antara dinosaurus dan burung. Fosil ini dianggap sebagai fosil transisi, karena menampilkan ciri-ciri dari kedua kelompok. Terungkap hanya dua tahun setelah karya penting Charles Darwin, On the Origin of Species (1859), penemuannya memicu perdebatan awal antara pendukung biologi evolusioner dan kreasionisme. Burung purba ini sangat mirip dinosaurus sehingga, tanpa cetakan bulu yang jelas di batuan sekitarnya, setidaknya satu spesimen sempat keliru dianggap sebagai theropoda kecil Compsognathus.[261] Sejak tahun 1990-an, sejumlah dinosaurus berbulu tambahan telah ditemukan, memberikan bukti yang lebih kuat tentang hubungan erat antara dinosaurus dan burung modern. Banyak dari spesimen ini digali di lagerstätten Biota Jehol.[254] Jika struktur mirip bulu memang tersebar luas di kalangan dinosaurus non-avian, kurangnya bukti fosil yang melimpah mungkin disebabkan oleh fakta bahwa fitur-fitur halus seperti kulit dan bulu jarang terawetkan oleh fosilisasi dan karenanya sering absen dari catatan fosil.[262]
Kerangka
suntingKarena bulu sering dikaitkan dengan burung, dinosaurus berbulu sering disebut-sebut sebagai mata rantai yang hilang antara burung dan dinosaurus. Namun, berbagai fitur kerangka yang juga dimiliki oleh kedua kelompok tersebut mewakili jalur bukti penting lainnya bagi para paleontolog. Bagian kerangka dengan kemiripan penting meliputi leher, pubis, pergelangan tangan (karpal semilunar), lengan dan gelang bahu, furcula (tulang garpu), dan tulang dada. Perbandingan kerangka burung dan dinosaurus melalui analisis kladistik memperkuat alasan adanya hubungan tersebut.[263]
Anatomi lunak
sunting
Dinosaurus pemakan daging yang besar memiliki sistem kantung udara yang kompleks serupa dengan yang ditemukan pada burung modern, menurut penyelidikan tahun 2005 yang dipimpin oleh Patrick M. O'Connor. Paru-paru dinosaurus theropoda (karnivora yang berjalan dengan dua kaki dan memiliki kaki mirip burung) kemungkinan memompa udara ke dalam kantung-kantung berongga di kerangka mereka, seperti halnya pada burung. "Apa yang dulunya secara formal dianggap unik bagi burung ternyata hadir dalam beberapa bentuk pada leluhur burung," kata O'Connor.[264][265] Pada tahun 2008, para ilmuwan mendeskripsikan Aerosteon riocoloradensis, yang kerangkanya memberikan bukti terkuat hingga saat ini mengenai dinosaurus dengan sistem pernapasan mirip burung. Pemindaian CT pada tulang fosil Aerosteon mengungkapkan bukti keberadaan kantung udara di dalam rongga tubuh hewan tersebut.[225][266]
Bukti perilaku
suntingFosil troodontidae Mei dan Sinornithoides menunjukkan bahwa beberapa dinosaurus tidur dengan kepala terselip di bawah lengan mereka.[267] Perilaku ini, yang mungkin membantu menjaga kehangatan kepala, juga merupakan karakteristik burung modern. Beberapa spesimen deinonychosauria dan oviraptorosauria juga ditemukan terawetkan di atas sarang mereka, kemungkinan sedang mengeram dengan cara yang mirip burung.[268] Rasio antara volume telur dan massa tubuh dewasa di antara dinosaurus-dinosaurus ini menunjukkan bahwa telur-telur tersebut terutama dierami oleh pejantan dan bahwa anak-anaknya sangat prekosial, mirip dengan banyak burung tanah modern.[269]
Beberapa dinosaurus diketahui menggunakan batu empedal seperti burung modern. Batu-batu ini ditelan oleh hewan untuk membantu pencernaan dan memecah makanan serta serat keras begitu masuk ke dalam perut. Ketika ditemukan berasosiasi dengan fosil, batu empedal disebut gastrolit.[270]
Kepunahan kelompok utama
suntingSemua dinosaurus non-avian dan sebagian besar garis keturunan burung[271] punah dalam sebuah peristiwa kepunahan massal, yang disebut peristiwa kepunahan Kapur–Paleogen (K-Pg), pada akhir periode Kapur. Di atas batas Kapur–Paleogen, yang telah ditentukan bertanggal 66,038 ± 0,025 juta tahun yang lalu,[272] fosil dinosaurus non-avian menghilang secara tiba-tiba; ketiadaan fosil dinosaurus secara historis digunakan untuk menetapkan batuan ke zaman Kenozoikum yang mengikutinya. Sifat peristiwa yang menyebabkan kepunahan massal ini telah dipelajari secara ekstensif sejak tahun 1970-an, yang mengarah pada pengembangan dua mekanisme yang diperkirakan memainkan peran utama: peristiwa tumbukan ekstraterestrial di Semenanjung Yucatán, bersamaan dengan vulkanisme banjir basal di India. Namun, mekanisme spesifik dari peristiwa kepunahan tersebut dan sejauh mana dampaknya terhadap dinosaurus masih menjadi area penelitian yang sedang berlangsung.[273] Bersama dengan dinosaurus, banyak kelompok hewan lain menjadi punah: pterosaurus, reptil laut seperti mosasaurus dan plesiosaurus, beberapa kelompok mamalia, amonit (moluska mirip nautilus), rudist (bivalvia pembentuk terumbu), dan berbagai kelompok plankton laut.[274][275] Secara keseluruhan, sekitar 47% genera dan 76% spesies di Bumi punah selama peristiwa kepunahan K-Pg.[276] Ukuran yang relatif besar dari sebagian besar dinosaurus dan rendahnya keanekaragaman spesies dinosaurus bertubuh kecil pada akhir Kapur mungkin berkontribusi terhadap kepunahan mereka;[277] kepunahan garis keturunan burung yang tidak bertahan hidup mungkin juga disebabkan oleh ketergantungan pada habitat hutan atau kurangnya adaptasi untuk memakan biji-bijian demi kelangsungan hidup.[278][279]
Keanekaragaman pra-kepunahan
suntingTepat sebelum peristiwa kepunahan K-Pg, jumlah spesies dinosaurus non-avian yang ada secara global diperkirakan antara 628 dan 1078.[280] Masih belum pasti apakah keanekaragaman dinosaurus mengalami penurunan bertahap sebelum peristiwa kepunahan K-Pg, atau apakah dinosaurus sebenarnya berkembang pesat sebelum kepunahan. Formasi batuan dari kala Maastrichtian, yang secara langsung mendahului kepunahan, ditemukan memiliki keanekaragaman yang lebih rendah daripada kala Campanian sebelumnya, yang mengarah pada pandangan umum tentang penurunan jangka panjang dalam keanekaragaman.[274][275][281] Namun, perbandingan ini tidak memperhitungkan potensi pengawetan yang bervariasi antar unit batuan atau tingkat eksplorasi dan penggalian yang berbeda.[273] Pada tahun 1984, Dale Russell melakukan analisis untuk memperhitungkan bias-bias ini, dan tidak menemukan bukti adanya penurunan;[282] analisis lain oleh David Fastovsky dan rekan-rekannya pada tahun 2004 bahkan menunjukkan bahwa keanekaragaman dinosaurus terus meningkat hingga kepunahan,[283] namun analisis ini telah dibantah.[284] Sejak saat itu, berbagai pendekatan berbeda berdasarkan statistik dan model matematika secara bervariasi mendukung baik kepunahan yang tiba-tiba[273][280][285] maupun penurunan bertahap.[286][287] Tren keanekaragaman pada akhir Kapur mungkin bervariasi antar garis keturunan dinosaurus: telah disarankan bahwa sauropoda tidak mengalami penurunan, sementara ornithischia dan theropoda sedang menurun.[288][289]
Peristiwa tumbukan
sunting

Hipotesis tumbukan bolide, yang pertama kali menarik perhatian luas pada tahun 1980 oleh Walter Alvarez, Luis Alvarez, dan rekan-rekannya, menghubungkan peristiwa kepunahan K-Pg dengan tumbukan bolide (proyektil ekstraterestrial).[290] Alvarez dan rekan-rekannya mengusulkan bahwa peningkatan mendadak pada kadar iridium, yang terekam di seluruh dunia dalam endapan batuan pada batas Kapur–Paleogen, adalah bukti langsung dari tumbukan tersebut.[291] Kuarsa terkejut, yang mengindikasikan gelombang kejut kuat yang memancar dari suatu tumbukan, juga ditemukan di seluruh dunia.[292] Lokasi tumbukan yang sebenarnya tetap sulit ditemukan sampai sebuah kawah berukuran lebar 180 km (110 mi) ditemukan di Semenanjung Yucatán di Meksiko tenggara, dan dipublikasikan dalam makalah tahun 1991 oleh Alan Hildebrand dan rekan-rekannya.[293] Sekarang, sebagian besar bukti menunjukkan bahwa sebuah bolide dengan lebar 5 hingga 15 kilometer (3 hingga 9+1⁄2 mil) menumbuk Semenanjung Yucatán 66 juta tahun yang lalu, membentuk kawah ini[294] dan menciptakan "mekanisme pemusnahan" yang memicu peristiwa kepunahan.[295][296][297]
Dalam hitungan jam, tumbukan Chicxulub akan menimbulkan efek langsung seperti gempa bumi,[298] tsunami,[299] dan badai api global yang kemungkinan besar membunuh hewan yang tidak berlindung dan memicu kebakaran hutan.[300][301] Namun, peristiwa ini juga akan memiliki konsekuensi jangka panjang bagi lingkungan. Dalam hitungan hari, aerosol sulfat yang dilepaskan dari batuan di lokasi tumbukan akan berkontribusi pada hujan asam dan pengasaman laut.[302][303] Aerosol jelaga diperkirakan telah menyebar ke seluruh dunia selama bulan-bulan dan tahun-tahun berikutnya; mereka akan mendinginkan permukaan Bumi dengan memantulkan radiasi termal, dan sangat memperlambat fotosintesis dengan menghalangi sinar matahari, sehingga menciptakan musim dingin tumbukan.[273][304][305] (Peran ini dikaitkan dengan aerosol sulfat sampai eksperimen menunjukkan sebaliknya.[303]) Terhentinya fotosintesis akan menyebabkan runtuhnya jaring-jaring makanan yang bergantung pada tanaman berdaun, yang mencakup semua dinosaurus kecuali burung pemakan biji-bijian.[279]
Trap Deccan
suntingPada saat kepunahan K-Pg, Deccan Traps yang merupakan banjir basal di India sedang meletus secara aktif. Letusan-letusan ini dapat dipisahkan menjadi tiga fase di sekitar batas K-Pg, dua sebelum batas tersebut dan satu setelahnya. Fase kedua, yang terjadi sangat dekat dengan batas tersebut, diperkirakan telah mengekstrusi 70 hingga 80% dari volume letusan ini dalam pulsa intermiten yang terjadi dengan selang waktu sekitar 100.000 tahun.[306][307] Gas rumah kaca seperti karbon dioksida dan belerang dioksida dilepaskan oleh aktivitas vulkanik ini,[308][309] yang mengakibatkan perubahan iklim melalui perturbasi suhu sekitar 3 °C (5,4 °F) tetapi mungkin setinggi 7 °C (13 °F).[310] Seperti halnya tumbukan Chicxulub, letusan ini mungkin juga melepaskan aerosol sulfat, yang akan menyebabkan hujan asam dan pendinginan global.[311] Namun, karena margin kesalahan yang besar dalam penanggalan letusan, peran Deccan Traps dalam kepunahan K-Pg masih belum jelas.[272][273][312]
Sebelum tahun 2000, argumen bahwa letusan Deccan Traps—sebagai lawan dari tumbukan Chicxulub—menyebabkan kepunahan biasanya dikaitkan dengan pandangan bahwa kepunahan tersebut terjadi secara bertahap. Sebelum penemuan kawah Chicxulub, Deccan Traps digunakan untuk menjelaskan lapisan iridium global;[308][313] bahkan setelah penemuan kawah tersebut, tumbukan itu masih dianggap hanya memiliki efek regional, bukan global, terhadap peristiwa kepunahan.[314] Sebagai tanggapan, Luis Alvarez menolak aktivitas vulkanik sebagai penjelasan untuk lapisan iridium dan kepunahan secara keseluruhan.[315] Namun, sejak saat itu, sebagian besar peneliti telah mengadopsi posisi yang lebih moderat, yang mengidentifikasi tumbukan Chicxulub sebagai penyebab utama kepunahan sekaligus mengakui bahwa Deccan Traps mungkin juga berperan. Walter Alvarez sendiri telah mengakui bahwa Deccan Traps dan faktor ekologi lainnya mungkin berkontribusi terhadap kepunahan selain tumbukan Chicxulub.[316] Beberapa perkiraan menempatkan awal fase kedua dalam letusan Deccan Traps dalam kurun waktu 50.000 tahun setelah tumbukan Chicxulub.[317] Dikombinasikan dengan pemodelan matematika dari gelombang seismik yang akan dihasilkan oleh tumbukan tersebut, hal ini mengarah pada dugaan bahwa tumbukan Chicxulub mungkin telah memicu letusan-letusan ini dengan meningkatkan permeabilitas plume mantel yang mendasari Deccan Traps.[318][319]
Apakah Deccan Traps merupakan penyebab utama kepunahan, setara dengan tumbukan Chicxulub, masih belum pasti. Para pendukung hipotesis ini menganggap dampak iklim dari belerang dioksida yang dilepaskan setara dengan dampak tumbukan Chicxulub, dan juga mencatat peran vulkanisme banjir basal dalam kepunahan massal lainnya seperti peristiwa kepunahan Perm–Trias.[320][321] Mereka menganggap tumbukan Chicxulub memperburuk perubahan iklim yang sedang berlangsung akibat letusan tersebut.[322] Sementara itu, para penentang menunjukkan sifat kepunahan yang tiba-tiba dan bahwa pulsa lain dalam aktivitas Deccan Traps dengan besaran yang sebanding tampaknya tidak menyebabkan kepunahan. Mereka juga berpendapat bahwa penyebab kepunahan massal yang berbeda harus dinilai secara terpisah.[323] Pada tahun 2020, Alfio Chiarenza dan rekan-rekannya menyarankan bahwa Deccan Traps bahkan mungkin memiliki efek sebaliknya: mereka mengusulkan bahwa pemanasan jangka panjang yang disebabkan oleh emisi karbon dioksidanya mungkin telah meredam musim dingin tumbukan dari hantaman Chicxulub.[297]
Kemungkinan penyintas Paleosen
suntingSisa-sisa dinosaurus non-avian kadang-kadang ditemukan di atas batas K-Pg. Pada tahun 2000, Spencer Lucas dan rekan-rekannya melaporkan penemuan sebuah tulang paha kanan hadrosauridae di Cekungan San Juan di New Mexico, dan mendeskripsikannya sebagai bukti adanya dinosaurus Paleosen. Unit batuan tempat tulang tersebut ditemukan diperkirakan berasal dari kala Paleosen awal, sekitar 64,8 juta tahun yang lalu.[324] Jika tulang tersebut tidak diendapkan kembali oleh proses pelapukan, hal itu akan memberikan bukti bahwa beberapa populasi dinosaurus bertahan hidup setidaknya setengah juta tahun hingga era Kenozoikum.[325] Bukti lain mencakup keberadaan sisa-sisa dinosaurus di Formasi Hell Creek hingga 13 m (43 ft) di atas batas Kapur–Paleogen , yang merepresentasikan selang waktu 40.000 tahun. Hal ini telah digunakan untuk mendukung pandangan bahwa kepunahan K-Pg terjadi secara bertahap.[326] Namun, dinosaurus yang diduga berasal dari Paleosen ini dianggap oleh banyak peneliti lain sebagai fosil rombakan (reworked fossil), yaitu fosil yang hanyut dari lokasi aslinya dan kemudian terkubur kembali dalam sedimen yang lebih muda.[327][328][329] Perkiraan usia tersebut juga dianggap tidak dapat diandalkan.[330]
Penggambaran budaya
sunting
Menurut standar manusia, dinosaurus adalah makhluk dengan penampilan yang fantastis dan ukuran yang sering kali sangat besar. Dengan demikian, mereka telah memikat imajinasi populer dan menjadi bagian abadi dari budaya manusia. Masuknya kata "dinosaurus" ke dalam bahasa sehari-hari umum mencerminkan pentingnya budaya hewan tersebut: dalam bahasa Inggris, "dinosaurus" (dinosaur) sering digunakan untuk menggambarkan segala sesuatu yang besar secara tidak praktis, usang, atau ditakdirkan untuk punah.[331]
Antusiasme publik terhadap dinosaurus pertama kali berkembang di Inggris pada era Victoria, di mana pada tahun 1854, tiga dekade setelah deskripsi ilmiah pertama tentang sisa-sisa dinosaurus, sekumpulan patung dinosaurus yang tampak hidup diresmikan di Taman Crystal Palace, London. Dinosaurus Crystal Palace terbukti sangat populer sehingga pasar replika berukuran kecil segera berkembang pesat. Pada dekade-dekade berikutnya, pameran dinosaurus dibuka di taman dan museum di seluruh dunia, memastikan bahwa generasi-generasi penerus akan diperkenalkan pada hewan-hewan ini dengan cara yang imersif dan mengasyikkan.[332] Popularitas dinosaurus yang abadi, pada gilirannya, telah menghasilkan pendanaan publik yang signifikan bagi sains dinosaurus, dan sering kali memacu penemuan-penemuan baru. Di Amerika Serikat, misalnya, persaingan antar-museum untuk menarik perhatian publik mengarah langsung pada Perang Tulang pada tahun 1880-an dan 1890-an, di mana sepasang paleontolog yang berseteru memberikan kontribusi ilmiah yang sangat besar.[333]
Minat populer terhadap dinosaurus telah mendorong kemunculan mereka dalam sastra, film, dan media lainnya. Dimulai pada tahun 1852 dengan penyebutan sekilas dalam Bleak House karya Charles Dickens,[334] dinosaurus telah ditampilkan dalam sejumlah besar karya fiksi. Novel Jules Verne tahun 1864 Journey to the Center of the Earth, buku Sir Arthur Conan Doyle tahun 1912 The Lost World, film animasi tahun 1914 Gertie the Dinosaur (yang menampilkan dinosaurus animasi pertama), film ikonik tahun 1933 King Kong, Godzilla tahun 1954 beserta banyak sekuelnya, novel terlaris tahun 1990 Jurassic Park karya Michael Crichton dan adaptasi filmnya tahun 1993 hanyalah beberapa contoh penting kemunculan dinosaurus dalam fiksi. Penulis karya nonfiksi minat umum tentang dinosaurus, termasuk beberapa paleontolog terkemuka, sering berusaha menggunakan hewan-hewan ini sebagai cara untuk mendidik pembaca tentang sains secara umum. Dinosaurus ada di mana-mana dalam periklanan; banyak perusahaan telah merujuk dinosaurus dalam iklan cetak atau televisi, baik untuk menjual produk mereka sendiri atau untuk mencirikan pesaing mereka sebagai lamban, bodoh, atau usang.[335][336]
Lihat pula
sunting- Pola makan dan cara makan dinosaurus
- Sejarah evolusi kehidupan
- Daftar unit stratigrafi yang mengandung dinosaurus
- Daftar genus dinosaurus
- Daftar genus burung
- Daftar burung
- Daftar dinosaurus yang dinamai secara informal
- Daftar film yang menampilkan dinosaurus
Catatan
sunting- ^ Dinosaurus (termasuk burung) adalah anggota kelompok alami Reptilia. Biologi dinosaurus tidak persis sesuai dengan kelas Reptilia yang usang dari taksonomi Linnaean, yang terdiri dari amniota berdarah dingin tanpa rambut atau bulu. Karena taksonomi Linnaean dirumuskan untuk hewan modern sebelum adanya penelitian evolusi dan paleontologi, sistem ini gagal memperhitungkan hewan punah dengan ciri perantara di antara kelas-kelas tradisional.
Referensi
sunting- ^ Matthew G. Baron; Megan E. Williams (2018). "A re-evaluation of the enigmatic dinosauriform Caseosaurus crosbyensis from the Late Triassic of Texas, USA and its implications for early dinosaur evolution". Acta Palaeontologica Polonica. 63. DOI:10.4202/app.00372.2017.
- ^ Andrea Cau (2018). "The assembly of the avian body plan: a 160-million-year long process" Diarsipkan December 3, 2024, di Wayback Machine. (PDF). Bollettino della Società Paleontologica Italiana. 57 (1): 1–25.
- ^ Ferigolo, Jorge; Langer, Max C. (2007). "A Late Triassic dinosauriform from south Brazil and the origin of the ornithischian predentary bone". Historical Biology. 19 (1): 23–33. Bibcode:2007HBio...19...23F. doi:10.1080/08912960600845767. ISSN 0891-2963. S2CID 85819339.
- ^ Langer, Max C.; Ferigolo, Jorge (2013). "The Late Triassic dinosauromorph Sacisaurus agudoensis (Caturrita Formation; Rio Grande do Sul, Brazil): anatomy and affinities". Geological Society, London, Special Publications (dalam bahasa Inggris). 379 (1): 353–392. Bibcode:2013GSLSP.379..353L. doi:10.1144/SP379.16. ISSN 0305-8719. S2CID 131414332. Diarsipkan dari versi aslinya tanggal August 14, 2021. Diakses tanggal March 26, 2021.
- ^ Cabreira, S.F.; Kellner, A.W.A.; Dias-da-Silva, S.; da Silva, L.R.; Bronzati, M.; de Almeida Marsola, J.C.; Müller, R.T.; de Souza Bittencourt, J.; Batista, B.J.; Raugust, T.; Carrilho, R.; Brodt, A.; Langer, M.C. (2016). "A Unique Late Triassic Dinosauromorph Assemblage Reveals Dinosaur Ancestral Anatomy and Diet". Current Biology. 26 (22): 3090–3095. Bibcode:2016CBio...26.3090C. doi:10.1016/j.cub.2016.09.040. ISSN 0960-9822. PMID 27839975.
- ^ Müller, Rodrigo Temp; Garcia, Maurício Silva (August 26, 2020). "A paraphyletic 'Silesauridae' as an alternative hypothesis for the initial radiation of ornithischian dinosaurs". Biology Letters. 16 (8) 20200417. doi:10.1098/rsbl.2020.0417. PMC 7480155. PMID 32842895.
- ^ a b c d Weishampel, Dodson & Osmólska 2004, hlm. 7–19, chpt. 1: "Origin and Relationships of Dinosauria" by Michael J. Benton.
- ^ Olshevsky 2000
- ^ a b Langer, Max C.; Ezcurra, Martin D.; Bittencourt, Jonathas S.; Novas, Fernando E. (February 2010). "The origin and early evolution of dinosaurs". Biological Reviews. 85 (1). Cambridge: Cambridge Philosophical Society: 65–66, 82. doi:10.1111/j.1469-185x.2009.00094.x. hdl:11336/103412. ISSN 1464-7931. PMID 19895605. S2CID 34530296.
- ^ "Using the tree for classification". Understanding Evolution. Berkeley, California: Universitas California. Diarsipkan dari versi aslinya tanggal 31 Agustus 2019. Diakses tanggal 14 Oktober 2019.
- ^ a b c d e f g Weishampel, Dodson & Osmólska 2004, hlm. 210–231, chpt. 11: "Basal Avialae" by Kevin Padian.
- ^ Wade, Nicholas (22 Maret 2017). "Shaking Up the Dinosaur Family Tree". The New York Times. New York. ISSN 0362-4331. Diarsipkan dari versi aslinya tanggal 7 April 2018. Diakses tanggal 30 Oktober 2019. "Versi artikel ini muncul di cetakan pada 28 Maret 2017, di Halaman D6 edisi New York dengan judul: Shaking Up the Dinosaur Family Tree."
- ^ Baron, Matthew G.; Norman, David B.; Barrett, Paul M. (2017). "A new hypothesis of dinosaur relationships and early dinosaur evolution". Nature. 543 (7646). London: Nature Research: 501–506. Bibcode:2017Natur.543..501B. doi:10.1038/nature21700. ISSN 0028-0836. PMID 28332513. S2CID 205254710.
- ^ Glut 1997, hlm. 40
- ^ Lambert & The Diagram Group 1990, hlm. 288
- ^ Farlow & Brett-Surman 1997, hlm. 607–624, chpt. 39: "Major Groups of Non-Dinosaurian Vertebrates of the Mesozoic Era" by Michael Morales.
- ^ a b Tennant, Jonathan P.; Chiarenza, Alfio Alessandro; Baron, Matthew (19 Februari 2018). "How has our knowledge of dinosaur diversity through geologic time changed through research history?". PeerJ. 6 e4417. doi:10.7717/peerj.4417. PMC 5822849. PMID 29479504.
- ^ Starrfelt, Jostein; Liow, Lee Hsiang (2016). "How many dinosaur species were there? Fossil bias and true richness estimated using a Poisson sampling model". Philosophical Transactions of the Royal Society B: Biological Sciences. 371 (1691) 20150219. doi:10.1098/rstb.2015.0219. PMC 4810813. PMID 26977060.
- ^ Wang, Steve C.; Dodson, Peter (2006). "Estimating the diversity of dinosaurs". Proc. Natl. Acad. Sci. U.S.A. 103 (37). Washington, D.C.: National Academy of Sciences: 13601–13605. Bibcode:2006PNAS..10313601W. doi:10.1073/pnas.0606028103. ISSN 0027-8424. PMC 1564218. PMID 16954187.
- ^ Russell, Dale A. (1995). "China and the lost worlds of the dinosaurian era". Historical Biology. 10 (1). Milton Park, Oxfordshire: Taylor & Francis: 3–12. Bibcode:1995HBio...10....3R. doi:10.1080/10292389509380510. ISSN 0891-2963.
- ^ Starrfelt, Jostein; Liow, Lee Hsiang (2016). "How many dinosaur species were there? Fossil bias and true richness estimated using a Poisson sampling model". Philosophical Transactions of the Royal Society B. 371 (1691) 20150219. London: Royal Society. doi:10.1098/rstb.2015.0219. ISSN 0962-8436. PMC 4810813. PMID 26977060.
- ^ Black, Riley (23 Maret 2016). "Most Dinosaur Species Are Still Undiscovered". National Geographic News. Diarsipkan dari versi aslinya tanggal 6 Maret 2021. Diakses tanggal 6 Juni 2021.
- ^ Gill, F.; Donsker, D.; Rasmussen, P. (2021). "Welcome". IOC World Bird List 11.1. Diarsipkan dari versi aslinya tanggal 30 Juli 2023. Diakses tanggal 25 Januari 2021.
- ^ MacLeod, Norman; Rawson, Peter F.; Forey, Peter L.; et al. (1997). "The Cretaceous–Tertiary biotic transition". Journal of the Geological Society. 154 (2). London: Geological Society of London: 265–292. Bibcode:1997JGSoc.154..265M. doi:10.1144/gsjgs.154.2.0265. ISSN 0016-7649. S2CID 129654916.
- ^ a b Amiot, Romain; Buffetaut, Éric; Lécuyer, Christophe; et al. (2010). "Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods". Geology. 38 (2). Boulder, CO: Geological Society of America: 139–142. Bibcode:2010Geo....38..139A. doi:10.1130/G30402.1. ISSN 0091-7613.
- ^ a b c Brusatte 2012, hlm. 9–20, 21
- ^ Nesbitt, Sterling J. (2011). "The Early Evolution of Archosaurs: Relationships and the Origin of Major Clades". Bulletin of the American Museum of Natural History. 2011 (352). New York: American Museum of Natural History: 1–292. doi:10.1206/352.1. hdl:2246/6112. ISSN 0003-0090. S2CID 83493714.
- ^ a b Paul 2000, hlm. 140–168, chpt. 3: "Classification and Evolution of the Dinosaur Groups" by Thomas R. Holtz Jr.
- ^ Smith, Dave; et al. "Dinosauria: Morphology". Berkeley: University of California Museum of Paleontology. Diarsipkan dari versi aslinya tanggal 12 November 2020. Diakses tanggal 16 Oktober 2019.
- ^ Langer, Max C.; Abdala, Fernando; Richter, Martha; Benton, Michael J. (1999). "Un dinosaure sauropodomorphe dans le Trias supérieur (Carnien) du Sud du Brésil" [A sauropodomorph dinosaur from the Upper Triassic (Carman) of southern Brazil]. Comptes Rendus de l'Académie des Sciences, Série IIA. 329 (7). Amsterdam: Elsevier on behalf of the French Academy of Sciences: 511–517. Bibcode:1999CRASE.329..511L. doi:10.1016/S1251-8050(00)80025-7. ISSN 1251-8050.
- ^ Nesbitt, Sterling J.; Irmis, Randall B.; Parker, William G. (2007). "A critical re-evaluation of the Late Triassic dinosaur taxa of North America". Journal of Systematic Palaeontology. 5 (2). Milton Park, Oxfordshire: Taylor & Francis on behalf of the Natural History Museum, London: 209–243. Bibcode:2007JSPal...5..209N. doi:10.1017/S1477201907002040. ISSN 1477-2019. S2CID 28782207.
- ^ This was recognized not later than 1909: Celeskey, Matt (2005). "Dr. W. J. Holland and the Sprawling Sauropods". The Hairy Museum of Natural History. Diarsipkan dari asli tanggal June 12, 2011. Diakses tanggal October 18, 2019.
- Holland, William J. (May 1910). "A Review of Some Recent Criticisms of the Restorations of Sauropod Dinosaurs Existing in the Museums of the United States, with Special Reference to that of Diplodocus Carnegiei in the Carnegie Museum". The American Naturalist. 44 (521). American Society of Naturalists: 259–283. Bibcode:1910ANat...44..258H. doi:10.1086/279138. ISSN 0003-0147. S2CID 84424110. Diakses tanggal October 18, 2019.
- The arguments and many of the images are also presented in Desmond 1975.
- ^ a b c Benton 2015, p. 205–210
- ^ Cowen 2005, hlm. 151–175, chpt. 12: "Dinosaurs".
- ^ a b Kubo, Tai; Benton, Michael J. (November 2007). "Evolution of hindlimb posture in archosaurs: limb stresses in extinct vertebrates" (PDF). Palaeontology. 50 (6). Hoboken, NJ: Wiley-Blackwell: 1519–1529. Bibcode:2007Palgy..50.1519K. doi:10.1111/j.1475-4983.2007.00723.x. ISSN 0031-0239. S2CID 140698705.
- ^ Dong 1992
- ^ "Dinosaur bones 'used as medicine'". BBC News. London: BBC. July 6, 2007. Diarsipkan dari versi aslinya tanggal August 27, 2019. Diakses tanggal November 4, 2019.
- ^ Paul 2000, hlm. 10–44, chpt. 1: "A Brief History of Dinosaur Paleontology" by Michael J. Benton.
- ^ a b Farlow & Brett-Surman 1997, hlm. 3–11, chpt. 1: "The Earliest Discoveries" by William A.S. Sarjeant.
- ^ Plot 1677, hlm. 131–139; illus. opp. p. 142, fig. 4
- ^ Plot 1677, hlm. [1]
- ^ "Robert Plot" (PDF). Learning more. Oxford: Oxford University Museum of Natural History. 2006. Diarsipkan dari asli (PDF) tanggal 1 Oktober 2006. Diakses tanggal 14 November 2019.
- ^ Lhuyd 1699, hlm. 67
- ^ Delair, Justin B.; Sarjeant, William A.S. (2002). "The earliest discoveries of dinosaurs: the records re-examined". Proceedings of the Geologists' Association. 113 (3). Amsterdam: Elsevier on behalf of the Geologists' Association: 185–197. Bibcode:2002PrGA..113..185D. doi:10.1016/S0016-7878(02)80022-0. ISSN 0016-7878.
- ^ Gunther 1968
- ^ Buckland, William (1824). "Notice on the Megalosaurus or great Fossil Lizard of Stonesfield". Transactions of the Geological Society of London. 1 (2). London: Geological Society of London: 390–396. doi:10.1144/transgslb.1.2.390. ISSN 2042-5295. S2CID 129920045. Diarsipkan (PDF) dari versi aslinya tanggal October 21, 2019. Diakses tanggal November 5, 2019.
- ^ Mantell, Gideon A. (1825). "Notice on the Iguanodon, a newly discovered fossil reptile, from the sandstone of Tilgate forest, in Sussex". Philosophical Transactions of the Royal Society of London. 115. London: Royal Society: 179–186. Bibcode:1825RSPT..115..179M. doi:10.1098/rstl.1825.0010. ISSN 0261-0523. JSTOR 107739.
- ^ Farlow & Brett-Surman 1997, hlm. 14, chpt. 2: "European Dinosaur Hunters" by Hans-Dieter Sues.
- ^ a b "The 'birth' of dinosaurs". More Than A Dodo. 28 April 2017. Diarsipkan dari versi aslinya tanggal 14 Maret 2023. Diakses tanggal 15 Maret 2023.
- ^ a b "The Birth of Dinosaurs: Richard Owen and Dinosauria". Biodiversity Heritage Library. 16 Oktober 2015. Diarsipkan dari versi aslinya tanggal 14 Maret 2023. Diakses tanggal 15 Maret 2023.
- ^ Brett-Surman, M. K.; Holtz, Thomas R.; Farlow, James O. (27 Juni 2012). The Complete Dinosaur. Indiana University Press. hlm. 25. ISBN 978-0-253-00849-7.
- ^ a b Owen 1842, p.103: "The combination of such characters ... will, it is presumed, be deemed sufficient ground for establishing a distinct tribe or sub-order of Saurian Reptiles, for which I would propose the name of Dinosauria*. (*Gr. δεινός, fearfully great; σαύρος, a lizard. ... )
- ^ "Dinosauria". Merriam-Webster Dictionary. Diakses tanggal 10 November 2019.
- ^ Crane, George R. (ed.). "Greek Dictionary Headword Search Results". Perseus 4.0. Medford and Somerville, MA: Tufts University. Diarsipkan dari versi aslinya tanggal December 10, 2019. Diakses tanggal October 13, 2019. Lemma for 'δεινός Diarsipkan December 10, 2019, di Wayback Machine.' from Henry George Liddell, Robert Scott, A Greek-English Lexicon (1940): 'fearful, terrible'.
- ^ Farlow & Brett-Surman 1997, hlm. ix–xi, Preface, "Dinosaurs: The Terrestrial Superlative" by James O. Farlow and M.K. Brett-Surman.
- ^ Rupke 1994
- ^ Prieto-Marquez, Albert; Weishampel, David B.; Horner, John R. (March 2006). "The dinosaur Hadrosaurus foulkii, from the Campanian of the East Coast of North America, with a reevaluation of the genus" (PDF). Acta Palaeontologica Polonica. 51 (1). Warsaw: Institute of Paleobiology, Polish Academy of Sciences: 77–98. ISSN 0567-7920. Diarsipkan (PDF) dari versi aslinya tanggal June 22, 2019. Diakses tanggal November 5, 2019.
- ^ Holmes 1998
- ^ a b c Taylor, M.P. (2010). "Sauropod dinosaur research: a historical review". Geological Society, London, Special Publications. 343 (1): 361–386. Bibcode:2010GSLSP.343..361T. doi:10.1144/SP343.22. S2CID 910635.
- ^ Naish, D. (2009). The Great Dinosaur Discoveries. London, UK: A & C Black Publishers Ltd. hlm. 89–93. ISBN 978-1-4081-1906-8.
- ^ Arbour, V. (2018). "Results roll in from the dinosaur renaissance". Science. 360 (6389): 611. Bibcode:2018Sci...360..611A. doi:10.1126/science.aat0451. S2CID 46887409.
- ^ a b Bakker, R.T. (1968). "The Superiority of Dinosaurs". Discovery: Magazine of the Peabody Museum of Natural History. 3 (2): 11–22. ISSN 0012-3625. OCLC 297237777.
- ^ a b Bakker, R.T. (1972). "Anatomical and Ecological Evidence of Endothermy in Dinosaurs". Nature. 238 (5359): 81–85. Bibcode:1972Natur.238...81B. doi:10.1038/238081a0. S2CID 4176132.
- ^ Bakker 1986
- ^ a b Benton, M.J. (2008). "Fossil quality and naming dinosaurs". Biology Letters. 4 (6): 729–732. Bibcode:2008BiLet...4..729B. doi:10.1098/rsbl.2008.0402. PMC 2614166. PMID 18796391.
- ^ a b Cashmore, D.D.; Mannion, P.D.; Upchurch, P.; Butler, R.J. (2020). "Ten more years of discovery: revisiting the quality of the sauropodomorph dinosaur fossil record". Palaeontology. 63 (6): 951–978. Bibcode:2020Palgy..63..951C. doi:10.1111/pala.12496. S2CID 219090716.
- ^ Cashmore, D.D.; Butler, R.J. (2019). "Skeletal completeness of the non-avian theropod dinosaur fossil record". Palaeontology. 62 (6): 951–981. Bibcode:2019Palgy..62..951C. doi:10.1111/pala.12436. S2CID 197571209.
- ^ Holtz, T.R. Jr.; Brett-Surman, M.K. (1997). "The Taxonomy and Systematics of Dinosaurs". The Complete Dinosaur. Bloomington: Indiana University Press. hlm. 209–223. ISBN 978-0-253-33349-0.
- ^ a b c St. Fleur, Nicholas (December 8, 2016). "That Thing With Feathers Trapped in Amber? It Was a Dinosaur Tail". Trilobites. The New York Times. New York. ISSN 0362-4331. Diarsipkan dari versi aslinya tanggal August 31, 2017. Diakses tanggal December 8, 2016.
- ^ Lockley, M.G.; Wright, J.L. (2000). "Reading About Dinosaurs – An Annotated Bibliography of Books". Journal of Geoscience Education. 48 (2): 167–178. Bibcode:2000JGeEd..48..167L. doi:10.5408/1089-9995-48.2.167. S2CID 151426669.
- ^ Lloyd, G.T.; Davis, K.E.; Pisani, D.; Tarver, J.E.; Ruta, R.; Sakamoto, M.; Hone, D.W.E.; Jennings, R.; Benton, M.J. (2008). "Dinosaurs and the Cretaceous Terrestrial Revolution". Proceedings of the Royal Society B. 275 (1650): 2483–2490. doi:10.1098/rspb.2008.0715. PMC 2603200. PMID 18647715.
- ^ a b Schweitzer, M.H. (2011). "Soft Tissue Preservation in Terrestrial Mesozoic Vertebrates". Annual Review of Earth and Planetary Sciences. 39: 187–216. Bibcode:2011AREPS..39..187S. doi:10.1146/annurev-earth-040610-133502.
- ^ a b Hooley, R.W. (1917). "II—On the Integument of Iguanodon bernissartensis, Boulenger, and of Morosaurus becklesii, Mantell". Geological Magazine. 4 (4): 148–150. Bibcode:1917GeoM....4..148H. doi:10.1017/s0016756800192386. S2CID 129640665. Diarsipkan dari versi aslinya tanggal 15 April 2023. Diakses tanggal 20 Februari 2023.
- ^ Osborn, H.F. (1912). "Integument of the iguanodont dinosaur Trachodon". Memoirs of the American Museum of Natural History. 1: 33–54.
- ^ Bell, P.R. (2014). "A review of hadrosaur skin impressions". Dalam Eberth, D.; Evans, D. (ed.). The Hadrosaurs: Proceedings of the International Hadrosaur Symposium. Bloomington: Princeton University Press. hlm. 572–590.
- ^ Eliason, C.M.; Hudson, L.; Watts, T.; Garza, H.; Clarke, J.A. (2017). "Exceptional preservation and the fossil record of tetrapod integument". Proceedings of the Royal Society B. 284 (1862): 1–10. doi:10.1098/rspb.2017.0556. PMC 5597822. PMID 28878057.
- ^ Benton, M.J. (1998). "Dinosaur fossils with soft parts" (PDF). Trends in Ecology & Evolution. 13 (8): 303–304. Bibcode:1998TEcoE..13..303B. doi:10.1016/s0169-5347(98)01420-7. PMID 21238317.
- ^ Zhou, Z.-H.; Wang, Y. (2017). "Vertebrate assemblages of the Jurassic Yanliao Biota and the Early Cretaceous Jehol Biota: Comparisons and implications". Palaeoworld. 26 (2): 241–252. Bibcode:2017Palae..26..241Z. doi:10.1016/j.palwor.2017.01.002.
- ^ Norell, M.A.; Xu, X. (2005). "Feathered Dinosaurs". Annual Review of Earth and Planetary Sciences. 33: 277–299. Bibcode:2005AREPS..33..277N. doi:10.1146/annurev.earth.33.092203.122511.
- ^ a b Roy, A.; Pittman, M.; Saitta, E.T.; Kaye, T.G.; Xu, X. (2020). "Recent advances in amniote palaeocolour reconstruction and a framework for future research". Biological Reviews. 95 (1): 22–50. doi:10.1111/brv.12552. PMC 7004074. PMID 31538399.
- ^ Vinther, J. (2020). "Reconstructing Vertebrate Paleocolor". Annual Review of Earth and Planetary Sciences. 48: 345–375. Bibcode:2020AREPS..48..345V. doi:10.1146/annurev-earth-073019-045641. S2CID 219768255.
- ^ Zhang, F.; Kearns, S.L.; Orr, P.J.; Benton, M.J.; Zhou, Z.; Johnson, D.; Xu, X.; Wang, X. (2010). "Fossilized melanosomes and the colour of Cretaceous dinosaurs and birds" (PDF). Nature. 463 (7284): 1075–1078. Bibcode:2010Natur.463.1075Z. doi:10.1038/nature08740. PMID 20107440. S2CID 205219587. Diarsipkan (PDF) dari versi aslinya tanggal 2 Februari 2014. Diakses tanggal 20 Februari 2023.
- ^ Smithwick, F.M.; Nicholls, R.; Cuthill, I.C.; Vinther, J. (2017). "Countershading and Stripes in the Theropod Dinosaur Sinosauropteryx Reveal Heterogeneous Habitats in the Early Cretaceous Jehol Biota". Current Biology. 27 (21): 3337–3343.e2. Bibcode:2017CBio...27E3337S. doi:10.1016/j.cub.2017.09.032. hdl:1983/8ee95b15-5793-42ad-8e57-da6524635349. PMID 29107548.
- ^ Vinther, J.; Nicholls, R.; Lautenschlager, S.; Pittman, M.; Kaye, T.G.; Rayfield, E.; Mayr, G.; Cuthill, I.C. (2016). "3D Camouflage in an Ornithischian Dinosaur". Current Biology. 26 (18): 2456–2462. Bibcode:2016CBio...26.2456V. doi:10.1016/j.cub.2016.06.065. PMC 5049543. PMID 27641767.
- ^ Lindgren, J.; Moyer, A.; Schweitzer, M.H.; Sjövall, P.; Uvdal, P.; Nilsson, D.E.; Heimdal, J.; Engdahl, A.; Gren, J.A.; Schultz, B.P.; Kear, B.P. (2015). "Interpreting melanin-based coloration through deep time: a critical review". Proceedings of the Royal Society B. 282 (1813) 20150614. Bibcode:2015PBioS.28250614L. doi:10.1098/rspb.2015.0614. PMC 4632609. PMID 26290071.
- ^ Schweitzer, M.H.; Lindgren, J.; Moyer, A.E. (2015). "Melanosomes and ancient coloration re-examined: a response to Vinther 2015 (DOI 10.1002/bies.201500018)". BioEssays. 37 (11): 1174–1183. doi:10.1002/bies.201500061. PMID 26434749. S2CID 45178498.
- ^ Zhou, Z. (2014). "The Jehol Biota, an Early Cretaceous terrestrial Lagerstätte: new discoveries and implications". National Science Review. 1 (4): 543–559. doi:10.1093/nsr/nwu055.
- ^ O'Connor, J.K.; Zhou, Z. (2019). "The evolution of the modern avian digestive system: insights from paravian fossils from the Yanliao and Jehol biotas". Palaeontology. 63 (1): 13–27. doi:10.1111/pala.12453. S2CID 210265348.
- ^ a b Dal Sasso, Cristiano; Signore, Marco (26 Maret 1998). "Exceptional soft-tissue preservation in a theropod dinosaur from Italy" (PDF). Nature. 392 (6674). London: Nature Research: 383–387. Bibcode:1998Natur.392..383D. doi:10.1038/32884. ISSN 0028-0836. S2CID 4325093. Diarsipkan (PDF) dari versi aslinya tanggal 20 September 2016.
- ^ Morell, V. (1993). "Dino DNA: the Hunt and the Hype". Science. 261 (5118): 160–162. Bibcode:1993Sci...261..160M. doi:10.1126/science.8327889. PMID 8327889.
- ^ Pawlicki, R.; Korbel, A.; Kubiak, H. (1996). "Cells, Collagen Fibrils and Vessels in Dinosaur Bone". Nature. 211 (5049): 655–657. doi:10.1038/211655a0. PMID 5968744. S2CID 4181847.
- ^ Pawlicki, R.; Nowogrodzka-Zagórska, M. (1998). "Blood vessels and red blood cells preserved in dinosaur bones". Annals of Anatomy - Anatomischer Anzeiger. 180 (1): 73–77. doi:10.1016/S0940-9602(98)80140-4. PMID 9488909.
- ^ a b Schweitzer, Mary H.; Wittmeyer, Jennifer L.; Horner, John R.; Toporski, Jan K. (2005). "Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex". Science. 307 (5717). Washington, D.C.: American Association for the Advancement of Science: 1952–1955. Bibcode:2005Sci...307.1952S. doi:10.1126/science.1108397. ISSN 0036-8075. PMID 15790853. S2CID 30456613.
- ^ Anderson, L.A. (2023). "A chemical framework for the preservation of fossil vertebrate cells and soft tissues". Earth-Science Reviews. 240 104367. Bibcode:2023ESRv..24004367A. doi:10.1016/j.earscirev.2023.104367. S2CID 257326012.
- ^ a b Schweitzer, M.H.; Zheng, W.; Organ, C.L.; Avci, R.; Suo, Z.; Freimark, L.M.; LeBleu, V.S.; Duncan, M.B.; van der Heiden, M.G.; Neveu, J.M.; Lane, W.S.; Cottrell, J.S.; Horner, J.R.; Cantley, L.C.; Kalluri, R.; Asara, J.M. (2009). "Biomolecular characterization and protein sequences of the Campanian hadrosaur B. canadensis". Science. 324 (5927): 626–631. Bibcode:2009Sci...324..626S. doi:10.1126/science.1165069. PMID 19407199. S2CID 5358680.
- ^ Organ, C.L.; Schweitzer, M.H.; Zheng, W.; Freimark, L.M.; Cantley, L.C.; Asara, J.M. (2008). "Molecular Phylogenetics of Mastodon and Tyrannosaurus rex". Science. 320 (5875): 499. Bibcode:2008Sci...320..499O. doi:10.1126/science.1154284. PMID 18436782. S2CID 24971064.
- ^ Schweitzer, M.H.; Zheng, W.; Cleland, T.P.; Bern, M. (2013). "Molecular analyses of dinosaur osteocytes support the presence of endogenous molecules". Bone. 52 (1). Amsterdam: Elsevier: 414–423. doi:10.1016/j.bone.2012.10.010. ISSN 8756-3282. PMID 23085295.
- ^ Bailleul, A.M.; Zheng, W.; Horner, J.R.; Hall, B.K.; Holliday, C.M.; Schweitzer, M.H. (2020). "Evidence of proteins, chromosomes and chemical markers of DNA in exceptionally preserved dinosaur cartilage". National Science Review. 7 (4): 815–822. doi:10.1093/nsr/nwz206. PMC 8289162. PMID 34692099.
- ^ Bertazzo, S.; Maidment, S.C.R.; Kallepitis, C.; et al. (2015). "Fibres and cellular structures preserved in 75-million-year-old dinosaur specimens". Nature Communications. 6 7352. Bibcode:2015NatCo...6.7352B. doi:10.1038/ncomms8352. ISSN 2041-1723. PMC 4468865. PMID 26056764.
- ^ Kaye, T.G.; Gaugler, G.; Sawlowicz, Z. (2008). "Dinosaurian Soft Tissues Interpreted as Bacterial Biofilms". PLOS ONE. 3 (7) e2808. Bibcode:2008PLoSO...3.2808K. doi:10.1371/journal.pone.0002808. PMC 2483347. PMID 18665236.
- ^ Peterson, J.E.; Lenczewski, M.E.; Scherer, R.P. (2010). "Influence of Microbial Biofilms on the Preservation of Primary Soft Tissue in Fossil and Extant Archosaurs". PLOS ONE. 5 (10) e13334. Bibcode:2010PLoSO...513334P. doi:10.1371/journal.pone.0013334. ISSN 1932-6203. PMC 2953520. PMID 20967227.
- ^ Buckley, M.; Warwood, S.; van Dongen, B.; Kitchener, A.C.; Manning, P.L. (2017). "A fossil protein chimera; difficulties in discriminating dinosaur peptide sequences from modern cross-contamination". Proceedings of the Royal Society B. 284 (1855). Bibcode:2017PBioS.28470544B. doi:10.1098/rspb.2017.0544. PMC 5454271. PMID 28566488.
- ^ Kump, Lee R.; Pavlov, Alexander; Arthur, Michael A. (2005). "Massive release of hydrogen sulfide to the surface ocean and atmosphere during intervals of oceanic anoxia". Geology. 33 (5). Boulder, CO: Geological Society of America: 397–400. Bibcode:2005Geo....33..397K. doi:10.1130/G21295.1. ISSN 0091-7613. S2CID 34821866.
- ^ Tanner, Lawrence H.; Lucas, Spencer G.; Chapman, Mary G. (March 2004). "Assessing the record and causes of Late Triassic extinctions" (PDF). Earth-Science Reviews. 65 (1–2). Amsterdam: Elsevier: 103–139. Bibcode:2004ESRv...65..103T. doi:10.1016/S0012-8252(03)00082-5. ISSN 0012-8252. Diarsipkan dari asli (PDF) tanggal October 25, 2007. Diakses tanggal October 22, 2007.
- ^ a b Griffin, C.T.; Wynd, B.M.; Munyikwa, D.; Broderick, T.J.; Zondo, M.; Tolan, S.; Langer, M.C.; Nesbitt, S.J.; Taruvinga, H.R. (2022). "Africa's oldest dinosaurs reveal early suppression of dinosaur distribution". Nature. 609 (7926): 313–319. Bibcode:2022Natur.609..313G. doi:10.1038/s41586-022-05133-x. ISSN 0028-0836. PMID 36045297. S2CID 251977824.
- ^ Desojo, J.B.; Fiorelli, L.E.; Ezcurra, M.D.; Martinelli, A.G.; Ramezani, J.; Da Rosa, A.A.S.; Belén von Baczko, M.; Jimena Trotteyn, M.; Montefeltro, F.C.; Ezpeleta, M.; Langer, M.C. (2020). "The Late Triassic Ischigualasto Formation at Cerro Las Lajas (La Rioja, Argentina): fossil tetrapods, high-resolution chronostratigraphy, and faunal correlations". Scientific Reports. 10 (1): 12782. Bibcode:2020NatSR..1012782D. doi:10.1038/s41598-020-67854-1. PMC 7391656. PMID 32728077.
- ^ Alcober, Oscar A.; Martinez, Ricardo N. (2010). "A new herrerasaurid (Dinosauria, Saurischia) from the Upper Triassic Ischigualasto Formation of northwestern Argentina". ZooKeys (63). Sofia: Pensoft Publishers: 55–81. Bibcode:2010ZooK...63...55A. doi:10.3897/zookeys.63.550. ISSN 1313-2989. PMC 3088398. PMID 21594020.
- ^ a b c Novas, F.E.; Agnolin, F.L.; Ezcurra, M.D.; Müller, R.T.; Martinelli, A.; Langer, M. (2021). "Review of the fossil record of early dinosaurs from South America, and its phylogenetic implications". Journal of South American Earth Sciences. 110 103341. Bibcode:2021JSAES.11003341N. doi:10.1016/j.jsames.2021.103341. ISSN 0895-9811.
- ^ Nesbitt, Sterling J; Sues, Hans-Dieter (2021). "The osteology of the early-diverging dinosaur Daemonosaurus chauliodus (Archosauria: Dinosauria) from the Coelophysis Quarry (Triassic: Rhaetian) of New Mexico and its relationships to other early dinosaurs". Zoological Journal of the Linnean Society. 191 (1): 150–179. doi:10.1093/zoolinnean/zlaa080.
- ^ a b Sereno, Paul C. (1999). "The Evolution of Dinosaurs". Science. 284 (5423). Washington, D.C.: American Association for the Advancement of Science: 2137–2147. doi:10.1126/science.284.5423.2137. ISSN 0036-8075. PMID 10381873. Diarsipkan (PDF) dari versi aslinya tanggal January 5, 2018. Diakses tanggal November 8, 2019.
- ^ Sereno, Paul C.; Forster, Catherine A.; Rogers, Raymond R.; Monetta, Alfredo M. (1993). "Primitive dinosaur skeleton from Argentina and the early evolution of Dinosauria". Nature. 361 (6407). London: Nature Research: 64–66. Bibcode:1993Natur.361...64S. doi:10.1038/361064a0. ISSN 0028-0836. S2CID 4270484.
- ^ a b Langer, Max C.; Ramezani, Jahandar; Da Rosa, Átila A.S. (May 2018). "U-Pb age constraints on dinosaur rise from south Brazil". Gondwana Research. 57. Amsterdam: Elsevier: 133–140. Bibcode:2018GondR..57..133L. doi:10.1016/j.gr.2018.01.005. ISSN 1342-937X.
- ^ Novas, F.E.; Ezcurra, M.D.; Chatterjee, S.; Kutty, T.S. (2011). "New dinosaur species from the Upper Triassic Upper Maleri and Lower Dharmaram formations of central India". Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 101 (3–4): 333–349. Bibcode:2010EESTR.101..333N. doi:10.1017/S1755691011020093. S2CID 128620874.
- ^ Marsicano, C.A.; Irmis, R.B.; Mancuso, A.C.; Mundil, R.; Chemale, F. (2016). "The precise temporal calibration of dinosaur origins". Proceedings of the National Academy of Sciences. 113 (3): 509–513. Bibcode:2016PNAS..113..509M. doi:10.1073/pnas.1512541112. PMC 4725541. PMID 26644579.
- ^ Nesbitt, Sterling J.; Barrett, Paul M.; Werning, Sarah; et al. (2012). "The oldest dinosaur? A Middle Triassic dinosauriform from Tanzania". Biology Letters. 9 (1) 20120949. London: Royal Society. doi:10.1098/rsbl.2012.0949. ISSN 1744-9561. PMC 3565515. PMID 23221875.
- ^ Marsicano, C.A.; Irmis, R.B.; Mancuso, A.C.; Mundil, R.; Chemale, F. (2015). "The precise temporal calibration of dinosaur origins". Proceedings of the National Academy of Sciences. 113 (3): 509–513. Bibcode:2016PNAS..113..509M. doi:10.1073/pnas.1512541112. ISSN 0027-8424. PMC 4725541. PMID 26644579.
- ^ Brusatte, Stephen L.; Benton, Michael J.; Ruta, Marcello; Lloyd, Graeme T. (2008). "Superiority, Competition, and Opportunism in the Evolutionary Radiation of Dinosaurs" (PDF). Science. 321 (5895). Washington, D.C.: American Association for the Advancement of Science: 1485–1488. Bibcode:2008Sci...321.1485B. doi:10.1126/science.1161833. hdl:20.500.11820/00556baf-6575-44d9-af39-bdd0b072ad2b. ISSN 0036-8075. PMID 18787166. S2CID 13393888. Diarsipkan (PDF) dari versi aslinya tanggal 2018-07-19. Diakses tanggal October 22, 2019.
- ^ Tanner, Spielmann & Lucas 2013, hlm. 562–566, "The first Norian (Revueltian) rhynchosaur: Bull Canyon Formation, New Mexico, U.S.A." by Justin A. Spielmann, Spencer G. Lucas and Adrian P. Hunt.
- ^ Sulej, Tomasz; Niedźwiedzki, Grzegorz (2019). "An elephant-sized Late Triassic synapsid with erect limbs". Science. 363 (6422). Washington, D.C.: American Association for the Advancement of Science: 78–80. Bibcode:2019Sci...363...78S. doi:10.1126/science.aal4853. ISSN 0036-8075. PMID 30467179. S2CID 53716186.
- ^ "Fossil tracks in the Alps help explain dinosaur evolution". Science and Technology. The Economist. London. April 19, 2018. ISSN 0013-0613. Diakses tanggal May 24, 2018.
- ^ a b c d Weishampel, Dodson & Osmólska 2004, hlm. 627–642, chpt. 27: "Mesozoic Biogeography of Dinosauria" by Thomas R. Holtz Jr., Ralph E. Chapman, and Matthew C. Lamanna.
- ^ a b c d e Weishampel, Dodson & Osmólska 2004, hlm. 614–626, chpt. 26: "Dinosaur Paleoecology" by David E. Fastovsky and Joshua B. Smith.
- ^ Sereno, Paul C.; Wilson, Jeffrey A.; Witmer, Lawrence M.; et al. (2007). Kemp, Tom (ed.). "Structural Extremes in a Cretaceous Dinosaur". PLOS ONE. 2 (11) e1230. San Francisco, CA: PLOS. Bibcode:2007PLoSO...2.1230S. doi:10.1371/journal.pone.0001230. ISSN 1932-6203. PMC 2077925. PMID 18030355.
- ^ Prasad, Vandana; Strömberg, Caroline A. E.; Alimohammadian, Habib; et al. (2005). "Dinosaur Coprolites and the Early Evolution of Grasses and Grazers". Science. 310 (5751). Washington, D.C.: American Association for the Advancement of Science: 1170–1180. Bibcode:2005Sci...310.1177P. doi:10.1126/science.1118806. ISSN 0036-8075. PMID 16293759. S2CID 1816461.
- ^ Weishampel, Dodson & Osmólska 2004, hlm. 672–684, chpt. 30: "Dinosaur Extinction" by J. David Archibald and David E. Fastovsky.
- ^ Dyke & Kaiser 2011, chpt. 14: "Bird Evolution Across the K–Pg Boundary and the Basal Neornithine Diversification" by Bent E. K. Lindow. DOI:10.1002/9781119990475.ch14
- ^ Cracraft, Joel (1968). "A Review of the Bathornithidae (Aves, Gruiformes), with Remarks on the Relationships of the Suborder Cariamae" (PDF). American Museum Novitates (2326). New York: American Museum of Natural History: 1–46. hdl:2246/2536. ISSN 0003-0082. Diakses tanggal October 22, 2019.
- ^ Alvarenga, Herculano; Jones, Washington W.; Rinderknecht, Andrés (May 2010). "The youngest record of phorusrhacid birds (Aves, Phorusrhacidae) from the late Pleistocene of Uruguay". Neues Jahrbuch für Geologie und Paläontologie. 256 (2). Stuttgart: E. Schweizerbart: 229–234. Bibcode:2010NJGPA.256..229A. doi:10.1127/0077-7749/2010/0052. ISSN 0077-7749. Diakses tanggal October 22, 2019.
- ^ Mayr 2009
- ^ Paul 1988, hlm. 248–250
- ^ Weishampel, Dodson & Osmólska 2004, hlm. 151–164, chpt. 7: "Therizinosauroidea" by James M. Clark, Teresa Maryańska, and Rinchen Barsbold.
- ^ Weishampel, Dodson & Osmólska 2004, hlm. 196–210, chpt. 10: "Dromaeosauridae" by Peter J. Makovicky and Mark A. Norell.
- ^ Taylor, Michael P.; Wedel, Mathew J. (2013). "Why sauropods had long necks; and why giraffes have short necks". PeerJ. 1 e36. Corte Madera, CA; London. doi:10.7717/peerj.36. ISSN 2167-8359. PMC 3628838. PMID 23638372.
- ^ a b c Alexander, R. McNeill (2006). "Dinosaur biomechanics". Proceedings of the Royal Society B. 273 (1596). London: Royal Society: 1849–1855. Bibcode:2006PBioS.273.1849A. doi:10.1098/rspb.2006.3532. ISSN 0962-8452. PMC 1634776. PMID 16822743.
- ^ Farlow, James O.; Dodson, Peter; Chinsamy, Anusuya (November 1995). "Dinosaur Biology". Annual Review of Ecology and Systematics. 26 (1). Palo Alto, CA: Annual Reviews: 445–471. Bibcode:1995AnRES..26..445F. doi:10.1146/annurev.es.26.110195.002305. ISSN 1545-2069.
- ^ Weishampel, Dodson & Osmólska 2004
- ^ Dodson & Gingerich 1993, hlm. 167–199, "On the rareness of big, fierce animals: speculations about the body sizes, population densities, and geographic ranges of predatory mammals and large carnivorous dinosaurs" by James O. Farlow.
- ^ Peczkis, Jan (1995). "Implications of body-mass estimates for dinosaurs". Journal of Vertebrate Paleontology. 14 (4). Milton Park, Oxfordshire: Taylor & Francis for the Society of Vertebrate Paleontology: 520–533. Bibcode:1995JVPal..14..520P. doi:10.1080/02724634.1995.10011575. ISSN 0272-4634. JSTOR 4523591.
- ^ "Dinosaur Evolution". Department of Paleobiology. Dinosaurs. Washington, D.C.: National Museum of Natural History. 2007. Diarsipkan dari asli tanggal November 11, 2007. Diakses tanggal November 21, 2007.
- ^ a b c Sander, P. Martin; Christian, Andreas; Clauss, Marcus; et al. (Februari 2011). "Biology of the sauropod dinosaurs: the evolution of gigantism". Biological Reviews. 86 (1). Cambridge: Cambridge Philosophical Society: 117–155. doi:10.1111/j.1469-185X.2010.00137.x. ISSN 1464-7931. PMC 3045712. PMID 21251189.
- ^ a b c Foster & Lucas 2006, hlm. 131–138, "Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus Cope, 1878" by Kenneth Carpenter.
- ^ Paul 2010
- ^ Colbert 1971
- ^ Mazzetta, Gerardo V.; Christiansenb, Per; Fariñaa, Richard A. (2004). "Giants and Bizarres: Body Size of Some Southern South American Cretaceous Dinosaurs" (PDF). Historical Biology. 16 (2–4). Milton Park, Oxfordshire: Taylor & Francis: 71–83. Bibcode:2004HBio...16...71M. CiteSeerX 10.1.1.694.1650. doi:10.1080/08912960410001715132. ISSN 0891-2963. S2CID 56028251. Diarsipkan (PDF) dari versi aslinya tanggal 25 Februari 2009.
- ^ Janensch, Werner (1950). "Die Skelettrekonstruktion von Brachiosaurus brancai" [The Skeleton Reconstruction of Brachiosaurus brancai] (PDF). Palaeontographica. Suplement VII (1. Reihe, Teil 3, Lieferung 2). Terjemahan oleh Gerhard Maier. Stuttgart: E. Schweizerbart: 97–103. OCLC 45923346. Diarsipkan (PDF) dari versi aslinya tanggal 11 Juli 2017. Diakses tanggal 24 Oktober 2019.
- ^ Lucas, Spencer G.; Herne, Matthew C.; Hecket, Andrew B.; et al. (2004). Reappraisal of Seismosaurus, a Late Jurassic Sauropod Dinosaur From New Mexico. 2004 Denver Annual Meeting (November 7–10, 2004). Vol. 36. Boulder, CO: Geological Society of America. hlm. 422. OCLC 62334058. Paper No. 181-4. Diarsipkan dari versi aslinya tanggal 8 Oktober 2019. Diakses tanggal 25 Oktober 2019.
- ^ Sellers, William Irvin.; Margetts, Lee; Coria, Rodolfo Aníbal; Manning, Phillip Lars (2013). Carrier, David (ed.). "March of the Titans: The Locomotor Capabilities of Sauropod Dinosaurs". PLOS ONE. 8 (10) e78733. San Francisco, CA: PLOS. Bibcode:2013PLoSO...878733S. doi:10.1371/journal.pone.0078733. ISSN 1932-6203. PMC 3864407. PMID 24348896.
- ^ Lovelace, David M.; Hartman, Scott A.; Wahl, William R. (October–December 2007). "Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny". Arquivos do Museu Nacional. 65 (4). Rio de Janeiro: National Museum of Brazil; Federal University of Rio de Janeiro: 527–544. CiteSeerX 10.1.1.603.7472. ISSN 0365-4508. Diakses tanggal October 26, 2019.
- ^ Carpenter, Kenneth (2018). "Maraapunisaurus fragillimus, N.G. (formerly Amphicoelias fragillimus), a basal Rebbachisaurid from the Morrison Formation (Upper Jurassic) of Colorado". Geology of the Intermountain West. 5: 227–244. doi:10.31711/giw.v5i0.28.
- ^ Paul, Gregory S. (2019). "Determining the largest known land animal: A critical comparison of differing methods for restoring the volume and mass of extinct animals" (PDF). Annals of the Carnegie Museum. 85 (4): 335–358. Bibcode:2019AnCM...85..335P. doi:10.2992/007.085.0403. S2CID 210840060. Diarsipkan (PDF) dari versi aslinya tanggal 28 Januari 2020. Diakses tanggal 2 Desember 2023.
- ^ Pal, Saurabh; Ayyasami, Krishnan (27 Juni 2022). "The lost titan of Cauvery". Geology Today (dalam bahasa Inggris). 38 (3): 112–116. Bibcode:2022GeolT..38..112P. doi:10.1111/gto.12390. ISSN 0266-6979. S2CID 250056201.
- ^ Paul, Gregory S.; Larramendi, Asier (11 April 2023). "Body mass estimate of Bruhathkayosaurus and other fragmentary sauropod remains suggest the largest land animals were about as big as the greatest whales". Lethaia (dalam bahasa Inggris). 56 (2): 1–11. Bibcode:2023Letha..56..2.5P. doi:10.18261/let.56.2.5. ISSN 0024-1164. S2CID 259782734.
- ^ Dal Sasso, Cristiano; Maganuco, Simone; Buffetaut, Éric; et al. (2005). "New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities" (PDF). Journal of Vertebrate Paleontology. 25 (4). Milton Park, Oxfordshire: Taylor & Francis for the Society of Vertebrate Paleontology: 888–896. doi:10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2. ISSN 0272-4634. S2CID 85702490. Diarsipkan dari asli (PDF) tanggal April 29, 2011. Diakses tanggal May 5, 2011.
- ^ a b Therrien, François; Henderson, Donald M. (2007). "My theropod is bigger than yours ... or not: estimating body size from skull length in theropods". Journal of Vertebrate Paleontology. 27 (1). Milton Park, Oxfordshire: Taylor & Francis for the Society of Vertebrate Paleontology: 108–115. doi:10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2. ISSN 0272-4634. S2CID 86025320.
- ^ Zhao, Xijin; Li, Dunjing; Han, Gang; et al. (2007). "Zhuchengosaurus maximus from Shandong Province". Acta Geoscientia Sinica. 28 (2). Beijing: Chinese Academy of Geological Sciences: 111–122. ISSN 1006-3021.
- ^ Weishampel, Dodson & Osmólska 2004, hlm. 438–463, chpt. 20: "Hadrosauridae" by John R. Horner David B. Weishampel, and Catherine A. Forster.
- ^ Norell, Gaffney & Dingus 2000
- ^ "Bee Hummingbird (Mellisuga helenae)". Birds.com. Paley Media. Diarsipkan dari versi aslinya tanggal 3 April 2015. Diakses tanggal 27 Oktober 2019.
- ^ a b Zhang, Fucheng; Zhou, Zhonghe; Xu, Xing; et al. (2008). "A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers". Nature. 455 (7216). London: Nature Research: 1105–1108. Bibcode:2008Natur.455.1105Z. doi:10.1038/nature07447. ISSN 0028-0836. PMID 18948955. S2CID 4362560.
- ^ a b Xu, Xing; Zhao, Qi; Norell, Mark; et al. (February 2008). "A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin". Chinese Science Bulletin. 54 (3). Amsterdam: Elsevier on behalf of Science in China Press: 430–435. Bibcode:2009SciBu..54..430X. doi:10.1007/s11434-009-0009-6. ISSN 1001-6538. S2CID 53445386.
- ^ Holtz 2007
- ^ Butler, Richard J.; Zhao, Qi (February 2009). "The small-bodied ornithischian dinosaurs Micropachycephalosaurus hongtuyanensis and Wannanosaurus yansiensis from the Late Cretaceous of China". Cretaceous Research. 30 (1). Amsterdam: Elsevier: 63–77. Bibcode:2009CrRes..30...63B. doi:10.1016/j.cretres.2008.03.002. ISSN 0195-6671.
- ^ Yans, Johan; Dejax, Jean; Pons, Denise; et al. (January–February 2005). "Implications paléontologiques et géodynamiques de la datation palynologique des sédiments à faciès wealdien de Bernissart (bassin de Mons, Belgique)" [Palaeontological and geodynamical implications of the palynological dating of the wealden facies sediments of Bernissart (Mons Basin, Belgium)]. Comptes Rendus Palevol (dalam bahasa Prancis). 4 (1–2). Amsterdam: Elsevier of behalf of the French Academy of Sciences: 135–150. Bibcode:2005CRPal...4..135Y. doi:10.1016/j.crpv.2004.12.003. ISSN 1631-0683.
- ^ Day, Julia J.; Upchurch, Paul; Norman, David B.; et al. (2002). "Sauropod Trackways, Evolution, and Behavior" (PDF). Science. 296 (5573). Washington, D.C.: American Association for the Advancement of Science: 1659. doi:10.1126/science.1070167. ISSN 0036-8075. PMID 12040187. S2CID 36530770.
- ^ Curry Rogers & Wilson 2005, hlm. 252–284, chpt. 9: "Steps in Understanding Sauropod Biology: The Importance of Sauropods Tracks" by Joanna L. Wright.
- ^ Varricchio, David J.; Sereno, Paul C.; Zhao, Xijin; et al. (2008). "Mud-trapped herd captures evidence of distinctive dinosaur sociality" (PDF). Acta Palaeontologica Polonica. 53 (4). Warsaw: Institute of Paleobiology, Polish Academy of Sciences: 567–578. Bibcode:2008AcPaP..53..567V. doi:10.4202/app.2008.0402. ISSN 0567-7920. S2CID 21736244. Diarsipkan (PDF) dari versi aslinya tanggal March 30, 2019. Diakses tanggal May 6, 2011.
- ^ Lessem & Glut 1993, hlm. 19–20, "Allosaurus"
- ^ Maxwell, W. Desmond; Ostrom, John H. (1995). "Taphonomy and paleobiological implications of Tenontosaurus–Deinonychus associations". Journal of Vertebrate Paleontology. 15 (4). Milton Park, Oxfordshire: Taylor & Francis for the Society of Vertebrate Paleontology: 707–712. Bibcode:1995JVPal..15..707M. doi:10.1080/02724634.1995.10011256. ISSN 0272-4634.
- ^ Roach, Brian T.; Brinkman, Daniel L. (April 2007). "A Reevaluation of Cooperative Pack Hunting and Gregariousness in Deinonychus antirrhopus and Other Nonavian Theropod Dinosaurs". Bulletin of the Peabody Museum of Natural History. 48 (1). New Haven, CT: Peabody Museum of Natural History: 103–138. doi:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2. ISSN 0079-032X. S2CID 84175628.
- ^ Tanke, Darren H. (1998). "Head-biting behavior in theropod dinosaurs: paleopathological evidence" (PDF). Gaia: Revista de Geociências (15). Lisbon: National Museum of Natural History and Science: 167–184. doi:10.7939/R34T6FJ1P. ISSN 0871-5424. S2CID 90552600. Diarsipkan dari asli (PDF) tanggal February 27, 2008.
- ^ "The Fighting Dinosaurs". New York: American Museum of Natural History. Diarsipkan dari asli tanggal January 18, 2012. Diakses tanggal December 5, 2007.
- ^ a b Carpenter, Kenneth (1998). "Evidence of predatory behavior by theropod dinosaurs" (PDF). Gaia: Revista de Geociências. 15. Lisbon: National Museum of Natural History and Science: 135–144. ISSN 0871-5424. Diarsipkan (PDF) dari versi aslinya tanggal 2013-09-26.
- ^ Rogers, Raymond R.; Krause, David W.; Curry Rogers, Kristina (2007). "Cannibalism in the Madagascan dinosaur Majungatholus atopus". Nature. 422 (6931). London: Nature Research: 515–518. Bibcode:2003Natur.422..515R. doi:10.1038/nature01532. ISSN 0028-0836. PMID 12673249. S2CID 4389583.
- ^ Schmitz, Lars; Motani, Ryosuke (2011). "Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology". Science. 332 (6030). Washington, D.C.: American Association for the Advancement of Science: 705–708. Bibcode:2011Sci...332..705S. doi:10.1126/science.1200043. ISSN 0036-8075. PMID 21493820. S2CID 33253407.
- ^ Varricchio, David J.; Martin, Anthony J.; Katsura, Yoshihiro (2007). "First trace and body fossil evidence of a burrowing, denning dinosaur". Proceedings of the Royal Society B. 274 (1616). London: Royal Society: 1361–1368. doi:10.1098/rspb.2006.0443. ISSN 0962-8452. PMC 2176205. PMID 17374596.
- ^ Chiappe & Witmer 2002
- ^ Chatterjee, Sankar; Templin, R. Jack (2007). "Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui" (PDF). Proc. Natl. Acad. Sci. U.S.A. 104 (5). Washington, D.C.: National Academy of Sciences: 1576–1580. Bibcode:2007PNAS..104.1576C. doi:10.1073/pnas.0609975104. ISSN 0027-8424. PMC 1780066. PMID 17242354. Diarsipkan (PDF) dari versi aslinya tanggal August 18, 2019. Diakses tanggal October 29, 2019.
- ^ Goriely, Alain; McMillen, Tyler (2002). "Shape of a Cracking Whip". Physical Review Letters. 88 (24) 244301. Ridge, NY: American Physical Society. Bibcode:2002PhRvL..88x4301G. doi:10.1103/PhysRevLett.88.244301. ISSN 0031-9007. PMID 12059302.
- ^ Henderson, Donald M. (2003). "Effects of stomach stones on the buoyancy and equilibrium of a floating crocodilian: a computational analysis". Canadian Journal of Zoology. 81 (8). Ottawa: NRC Research Press: 1346–1357. Bibcode:2003CaJZ...81.1346H. doi:10.1139/z03-122. ISSN 0008-4301.
- ^ a b Senter, Phil (30 June 2009). "Voices of the past: a review of Paleozoic and Mesozoic animal sounds". Historical Biology. 20 (4): 255–287. Bibcode:2008HBio...20..255S. doi:10.1080/08912960903033327. ISSN 0891-2963. S2CID 84473967.
- ^ Li, Quanguo; Gao, Ke-Qin; Vinther, Jakob; et al. (2010). "Plumage Color Patterns of an Extinct Dinosaur" (PDF). Science. 327 (5971). Washington, D.C.: American Association for the Advancement of Science: 1369–1372. Bibcode:2010Sci...327.1369L. doi:10.1126/science.1186290. ISSN 0036-8075. PMID 20133521. S2CID 206525132. Diarsipkan (PDF) dari versi aslinya tanggal March 30, 2019. Diakses tanggal November 7, 2019.
- ^ Yoshida, Junki; Kobayashi, Yoshitsugu; Norell, Mark A. (2023-02-15). "An ankylosaur larynx provides insights for bird-like vocalization in non-avian dinosaurs". Communications Biology (dalam bahasa Inggris). 6 (1): 152. doi:10.1038/s42003-023-04513-x. ISSN 2399-3642. PMC 9932143. PMID 36792659.
- ^ Yang, Yunfeng; King, James L.; Xu, Xing (2025). "A new neornithischian dinosaur from the Upper Jurassic Tiaojishan Formation of northern China". PeerJ. 13 e19664. doi:10.7717/peerj.19664. PMC 12258161. PMID 40661901.
- ^ Riede, Tobias; Eliason, Chad M.; Miller, Edward H.; et al. (2016). "Coos, booms, and hoots: the evolution of closed-mouth vocal behavior in birds". Evolution. 70 (8). Hoboken, NJ: John Wiley & Sons for the Society for the Study of Evolution: 1734–1746. Bibcode:2016Evolu..70.1734R. doi:10.1111/evo.12988. ISSN 0014-3820. PMID 27345722. S2CID 11986423.
- ^ Weishampel, David B. (Spring 1981). "Acoustic Analysis of Vocalization of Lambeosaurine Dinosaurs (Reptilia: Ornithischia)" (PDF). Paleobiology. 7 (2). Bethesda, MD: Paleontological Society: 252–261. doi:10.1017/S0094837300004036. ISSN 0094-8373. JSTOR 2400478. S2CID 89109302. Diarsipkan dari asli (PDF) tanggal October 6, 2014. Diakses tanggal October 30, 2019.
- ^ Miyashita, Tetsuto; Arbour, Victoria M.; Witmer, Lawrence M.; et al. (December 2011). "The internal cranial morphology of an armoured dinosaur Euoplocephalus corroborated by X-ray computed tomographic reconstruction" (PDF). Journal of Anatomy. 219 (6). Hoboken, NJ: John Wiley & Sons: 661–675. doi:10.1111/j.1469-7580.2011.01427.x. ISSN 1469-7580. PMC 3237876. PMID 21954840. Diarsipkan dari asli (PDF) tanggal September 24, 2015. Diakses tanggal October 30, 2019.
- ^ Riede, T. (2019). "The evolution of the syrinx: an acoustic theory". PLOS ONE. 17 (2) e2006507. doi:10.1371/journal.pbio.2006507. PMC 6366696. PMID 30730882.
- ^ Clarke, Julia A.; Chatterjee, Sankar; Zhiheng, Li; et al. (2016). "Fossil evidence of the avian vocal organ from the Mesozoic". Nature. 538 (7626). London: Nature Research: 502–505. Bibcode:2016Natur.538..502C. doi:10.1038/nature19852. hdl:11336/64786. ISSN 0028-0836. PMID 27732575. S2CID 4389926.
- ^ Kingsley, E.P.; et al. (2018). "Identity and novelty in the avian syrinx". Proceedings of the National Academy of Sciences of the United States of America. 115 (41): 10109–10217. Bibcode:2018PNAS..11510209K. doi:10.1073/pnas.1804586115. PMC 6187200. PMID 30249637.
- ^ Hansell 2000
- ^ a b Varricchio, David J.; Horner, John R.; Jackson, Frankie D. (2002). "Embryos and eggs for the Cretaceous theropod dinosaur Troodon formosus". Journal of Vertebrate Paleontology. 22 (3). Milton Park, Oxfordshire: Taylor & Francis for the Society of Vertebrate Paleontology: 564–576. doi:10.1671/0272-4634(2002)022[0564:EAEFTC]2.0.CO;2. ISSN 0272-4634. S2CID 85728452.
- ^ Lee, Andrew H.; Werning, Sarah (2008). "Sexual maturity in growing dinosaurs does not fit reptilian growth models". Proc. Natl. Acad. Sci. U.S.A. 105 (2). Washington, D.C.: National Academy of Sciences: 582–587. Bibcode:2008PNAS..105..582L. doi:10.1073/pnas.0708903105. ISSN 0027-8424. PMC 2206579. PMID 18195356.
- ^ Horner, John R.; Makela, Robert (1979). "Nest of juveniles provides evidence of family structure among dinosaurs". Nature. 282 (5736). London: Nature Research: 296–298. Bibcode:1979Natur.282..296H. doi:10.1038/282296a0. ISSN 0028-0836. S2CID 4370793.
- ^ "Discovering Dinosaur Behavior: 1960–present view". Encyclopædia Britannica. Chicago, IL: Encyclopædia Britannica, Inc. Diarsipkan dari asli tanggal December 13, 2013. Diakses tanggal October 30, 2019.
- ^ Currie et al. 2004, hlm. 234–250, chpt. 11: "Dinosaur Brooding Behavior and the Origin of Flight Feathers" by Thomas P. Hopp and Mark J. Orsen.
- ^ Reisz, Robert R.; Scott, Diane; Sues, Hans-Dieter; et al. (2005). "Embryos of an Early Jurassic Prosauropod Dinosaur and Their Evolutionary Significance" (PDF). Science. 309 (5735). Washington, D.C.: American Association for the Advancement of Science: 761–764. Bibcode:2005Sci...309..761R. doi:10.1126/science.1114942. ISSN 0036-8075. PMID 16051793. S2CID 37548361. Diarsipkan (PDF) dari versi aslinya tanggal 2018-07-22.
- ^ Clark, Neil D. L.; Booth, Paul; Booth, Claire L.; et al. (2004). "Dinosaur footprints from the Duntulm Formation (Bathonian, Jurassic) of the Isle of Skye" (PDF). Scottish Journal of Geology. 40 (1). London: Geological Society of London: 13–21. Bibcode:2004ScJG...40...13C. doi:10.1144/sjg40010013. ISSN 0036-9276. S2CID 128544813. Diarsipkan (PDF) dari versi aslinya tanggal July 22, 2013. Diakses tanggal December 12, 2019.
- ^ Zhou, Zhonghe; Zhang, Fucheng (2004). "A Precocial Avian Embryo from the Lower Cretaceous of China". Science. 306 (5696). Washington, D.C.: American Association for the Advancement of Science: 653. Bibcode:2004Sci...306..653Z. doi:10.1126/science.1100000. ISSN 0036-8075. PMID 15499011. S2CID 34504916.
- ^ Naish, Darren (May 15, 2012). "A drowned nesting colony of Late Cretaceous birds". Science. 306 (5696). Scientific American: 653. Bibcode:2004Sci...306..653Z. doi:10.1126/science.1100000. PMID 15499011. S2CID 34504916. Diarsipkan dari versi aslinya tanggal September 25, 2018. Diakses tanggal November 16, 2019.
- ^ Fernández, Mariela S.; García, Rodolfo A.; Fiorelli, Lucas; et al. (2013). "A Large Accumulation of Avian Eggs from the Late Cretaceous of Patagonia (Argentina) Reveals a Novel Nesting Strategy in Mesozoic Birds". PLOS ONE. 8 (4) e61030. San Francisco, CA: PLOS. Bibcode:2013PLoSO...861030F. doi:10.1371/journal.pone.0061030. ISSN 1932-6203. PMC 3629076. PMID 23613776.
- ^ Deeming, Denis Charles; Mayr, Gerald (May 2018). "Pelvis morphology suggests that early Mesozoic birds were too heavy to contact incubate their eggs" (PDF). Journal of Evolutionary Biology. 31 (5). Hoboken, NJ: Wiley-Blackwell on behalf of the European Society for Evolutionary Biology: 701–709. doi:10.1111/jeb.13256. ISSN 1010-061X. PMID 29485191. S2CID 3588317. Diarsipkan (PDF) dari versi aslinya tanggal 2020-06-02.
- ^ Myers, Timothy S.; Fiorillo, Anthony R. (2009). "Evidence for gregarious behavior and age segregation in sauropod dinosaurs" (PDF). Palaeogeography, Palaeoclimatology, Palaeoecology. 274 (1–2). Amsterdam: Elsevier: 96–104. Bibcode:2009PPP...274...96M. doi:10.1016/j.palaeo.2009.01.002. ISSN 0031-0182. Diarsipkan (PDF) dari versi aslinya tanggal 2020-05-29.
- ^ Vinther, Jakob; Nicholls, Robert; Kelly, Diane A. (February 22, 2021). "A cloacal opening in a non-avian dinosaur". Current Biology. 31 (4). Elsevier: R1 – R3. Bibcode:2021CBio...31.R182V. doi:10.1016/j.cub.2020.12.039. PMID 33472049. S2CID 231644183.
- ^ Weishampel, Dodson & Osmólska 2004, hlm. 643–659, chpt. 28: "Physiology of Nonavian Dinosaurs" by Anusuya Chinsamy and Willem J. Hillenius.
- ^ Pontzer, H.; Allen, V.; Hutchinson, J.R. (2009). "Biomechanics of running indicates endothermy in bipedal dinosaurs". PLOS ONE. 4 (11) e7783. Bibcode:2009PLoSO...4.7783P. doi:10.1371/journal.pone.0007783. ISSN 1932-6203. PMC 2772121. PMID 19911059.
- ^ a b Benson, R.B.J. (2018). "Dinosaur Macroevolution and Macroecology". Annual Review of Ecology, Evolution, and Systematics. 49: 379–408. doi:10.1146/annurev-ecolsys-110617-062231. S2CID 92837486.
- ^ Grady, J.M.; Enquist, B.J.; Dettweiler-Robinson, E.; Wright, N.A.; Smith, F.A. (2014). "Evidence for mesothermy in dinosaurs". Science. 344 (6189): 1268–1272. Bibcode:2014Sci...344.1268G. doi:10.1126/science.1253143. PMID 24926017. S2CID 9806780.
- ^ Legendre, L.J.; Guénard, G.; Botha-Brink, J.; Cubo, J. (2016). "Palaeohistological Evidence for Ancestral High Metabolic Rate in Archosaurs". Systematic Biology. 65 (6): 989–996. doi:10.1093/sysbio/syw033. PMID 27073251.
- ^ Seymour, R.S.; Bennett-Stamper, C.L.; Johnston, S.D.; Carrier, D.R.; Grigg, G.C. (2004). "Evidence for endothermic ancestors of crocodiles at the stem of archosaur evolution". Physiological and Biochemical Zoology. 77 (6): 1051–1067. doi:10.1093/sysbio/syw033. PMID 27073251.
- ^ Parsons 2001, hlm. 22–48, "The Heresies of Dr. Bakker".
- ^ Erickson, G.M. (2014). "On dinosaur growth". Annual Review of Earth and Planetary Sciences. 42 (1): 675–697. Bibcode:2014AREPS..42..675E. doi:10.1146/annurev-earth-060313-054858.
- ^ a b Bailleul, A.M.; O'Connor, J.; Schweitzer, M.H. (2019). "Dinosaur paleohistology: review, trends and new avenues of investigation". PeerJ. 7 e7764. Bibcode:2019PeerJ...7e7764B. doi:10.7717/peerj.7764. PMC 6768056. PMID 31579624.
- ^ De Ricqlès, A. (1974). "Evolution of endothermy: histological evidence" (PDF). Evolutionary Theory. 1 (2): 51–80. Diarsipkan (PDF) dari versi aslinya tanggal 17 April 2021.
- ^ De Ricqlès, A. (1980). "Tissue structures of dinosaur bone, functional significance and possible relation to dinosaur physiology". Dalam Thomas, R.D.K.; Olson, E.C. (ed.). A Cold Look at the Warm-Blooded Dinosaurs. New York: American Association for the Advancement of Science. hlm. 103–139.
- ^ Padian, K.; Horner, J.R.; de Ricqlès, A. (2004). "Growth in small dinosaurs and pterosaurs: the evolution of archosaurian growth strategies" (PDF). Journal of Vertebrate Paleontology. 24 (3): 555–571. doi:10.1671/0272-4634(2004)024[0555:GISDAP]2.0.CO;2. S2CID 86019906. Diarsipkan (PDF) dari versi aslinya tanggal 4 Februari 2023. Diakses tanggal 3 Januari 2023.
- ^ de Souza, G.A.; Bento Soares, M.; Souza Brum, A.; Zucolotto, M.; Sayão, J.M.; Carlos Weinschütz, L.; Kellner, A.W.A. (2020). "Osteohistology and growth dynamics of the Brazilian noasaurid Vespersaurus paranaensis Langer et al., 2019 (Theropoda: Abelisauroidea)". PeerJ. 8 e9771. doi:10.7717/peerj.9771. PMC 7500327. PMID 32983636. S2CID 221906765.
- ^ Untuk contoh pekerjaan ini yang dilakukan pada garis keturunan dinosaurus yang berbeda, lihat
- Erickson, G.M.; Tumanova, T.A. (2000). "Growth curve of Psittacosaurus mongoliensis Osborn (Ceratopsia: Psittacosauridae) inferred from long bone histology". Zoological Journal of the Linnean Society. 130 (4): 551–566. doi:10.1111/j.1096-3642.2000.tb02201.x. S2CID 84241148.
- Erickson, G.; Rogers, K.; Yerby, S. (2001). "Dinosaurian growth patterns and rapid avian growth rates". Nature. 412 (429–433): 429–433. Bibcode:2001Natur.412..429E. doi:10.1038/35086558. PMID 11473315. S2CID 4319534.Templat:Erratum
- Erickson, G.; Makovicky, P.; Currie, P.; Norell, M.A.; Yerby, S.A.; Brochu, C.A. (2004). "Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs" (PDF). Nature. 430 (7001): 772–775. Bibcode:2004Natur.430..772E. doi:10.1038/nature02699. PMID 15306807. S2CID 4404887. Diarsipkan (PDF) dari versi aslinya tanggal 14 Juli 2020.Templat:Erratum
- Lehman, T.M.; Woodward, H.N. (2008). "Modeling growth rates for sauropod dinosaurs" (PDF). Paleobiology. 34 (2): 264–281. doi:10.1666/0094-8373(2008)034[0264:MGRFSD]2.0.CO;2. S2CID 84163725. Diarsipkan (PDF) dari versi aslinya tanggal 4 Februari 2023. Diakses tanggal 3 Januari 2023.
- Horner, J.R.; de Ricqles, A.; Padian, K.; Scheetz, R.D. (2009). "Comparative long bone histology and growth of the "hypsilophodontid" dinosaurs Orodromeus makelai, Dryosaurus altus, and Tenontosaurus tillettii (Ornithischia: Euornithopoda)". Journal of Vertebrate Paleontology. 29 (3): 734–747. Bibcode:2009JVPal..29..734H. doi:10.1671/039.029.0312. S2CID 86277619.
- Woodward, H.; Freedman Fowler, E.; Farlow, J.; Horner, J. (2015). "Maiasaura, a model organism for extinct vertebrate population biology: A large sample statistical assessment of growth dynamics and survivorship". Paleobiology. 41 (4): 503–527. Bibcode:2015Pbio...41..503W. doi:10.1017/pab.2015.19. S2CID 85902880.
- ^ Amiot, R.; Lécuyer, C.; Buffetaut, E.; Escarguel, G.; Fluteau, F.; Martineau, F. (2006). "Oxygen isotopes from biogenic apatites suggest widespread endothermy in Cretaceous dinosaurs" (PDF). Earth and Planetary Science Letters. 246 (1–2): 41–54. Bibcode:2006E&PSL.246...41A. doi:10.1016/j.epsl.2006.04.018.
- ^ Amiot, R.; Wang, X.; Lécuyer, C.; Buffetaut, E.; Boudad, L.; Cavin, L.; Ding, Z.; Fluteau, F.; Kellner, A.W.A.; Tong, H.; Zhang, F. (2010). "Oxygen and carbon isotope compositions of middle Cretaceous vertebrates from North Africa and Brazil: ecological and environmental significance". Palaeogeography, Palaeoclimatology, Palaeoecology. 297 (2): 439–451. Bibcode:2010PPP...297..439A. doi:10.1016/j.palaeo.2010.08.027.
- ^ Kolodny, Y.; Luz, B.; Sander, M.; Clemens, W.A. (1996). "Dinosaur bones: fossils or pseudomorphs? The pitfalls of physiology reconstruction from apatitic fossils" (PDF). Palaeogeography, Palaeoclimatology, Palaeoecology. 126 (1–2): 161–171. Bibcode:1996PPP...126..161K. doi:10.1016/S0031-0182(96)00112-5. Diarsipkan (PDF) dari versi aslinya tanggal 4 Februari 2023. Diakses tanggal 3 Januari 2023.
- ^ Paul, G.S. (1988). "Physiological, migratorial, climatological, geophysical, survival, and evolutionary implications of Cretaceous polar dinosaurs". Journal of Paleontology. 62 (4): 640–652. JSTOR 1305468.
- ^ Clemens, W.A.; Nelms, L.G. (1993). "Paleoecological implications of Alaskan terrestrial vertebrate fauna in latest Cretaceous time at high paleolatitudes". Geology. 21 (6): 503–506. Bibcode:1993Geo....21..503C. doi:10.1130/0091-7613(1993)021<0503:PIOATV>2.3.CO;2.
- ^ Rich, T.H.; Vickers-Rich, P.; Gangloff, R.A. (2002). "Polar dinosaurs". Science. 295 (5557): 979–980. doi:10.1126/science.1068920. PMID 11834803. S2CID 28065814.
- ^ Buffetaut, E. (2004). "Polar dinosaurs and the question of dinosaur extinction: a brief review" (PDF). Palaeogeography, Palaeoclimatology, Palaeoecology. 214 (3): 225–231. doi:10.1016/j.palaeo.2004.02.050. Diarsipkan (PDF) dari versi aslinya tanggal 8 Juni 2020.
- ^ a b Sereno, Paul C.; Martinez, Ricardo N.; Wilson, Jeffrey A.; et al. (September 2008). Kemp, Tom (ed.). "Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina". PLOS ONE. 3 (9) e3303. San Francisco, CA: PLOS. Bibcode:2008PLoSO...3.3303S. doi:10.1371/journal.pone.0003303. ISSN 1932-6203. PMC 2553519. PMID 18825273.
- ^ O'Connor, P.M. (2009). "Evolution of archosaurian body plans: skeletal adaptations of an air-sac-based breathing apparatus in birds and other archosaurs". Journal of Experimental Zoology Part A: Ecological Genetics and Physiology. 311 (8): 629–646. Bibcode:2009JEZA..311..629O. doi:10.1002/jez.548. PMID 19492308.
- ^ Eagle, R.A.; Tütken, T.; Martin, T.S.; Tripati, A.K.; Fricke, H.C.; Connely, M.; Cifelli, R.L.; Eiler, J.M. (2011). "Dinosaur body temperatures determined from isotopic (13C-18O) ordering in fossil biominerals". Science. 333 (6041): 443–445. Bibcode:2011Sci...333..443E. doi:10.1126/science.1206196. PMID 21700837. S2CID 206534244.
- ^ Wedel, M.J. (2003). "Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs" (PDF). Paleobiology. 29 (2): 243–255. Bibcode:2003Pbio...29..243W. doi:10.1017/S0094837300018091. Diarsipkan (PDF) dari versi aslinya tanggal 18 Mei 2023. Diakses tanggal 3 Januari 2024.
- ^ Perry, S.F.; Christian, A.; Breuer, T.; Pajor, N.; Codd, J.R. (2009). "Implications of an avian-style respiratory system for gigantism in sauropod dinosaurs". Journal of Experimental Zoology Part A: Ecological Genetics and Physiology. 311 (8): 600–610. Bibcode:2009JEZA..311..600P. doi:10.1002/jez.517. PMID 19189317.
- ^ Alexander, R.M. (1998). "All-time giants: the largest animals and their problems". Palaeontology. 41: 1231–1245. Diarsipkan dari versi aslinya tanggal 27 September 2016. Diakses tanggal 2 Januari 2021.
- ^ Tsahar, E.; Martínez del Rio, C.; Izhaki, I.; Arad, Z. (2005). "Can birds be ammonotelic? Nitrogen balance and excretion in two frugivores" (PDF). The Journal of Experimental Biology. 208 (6): 1025–1034. Bibcode:2005JExpB.208.1025T. doi:10.1242/jeb.01495. ISSN 0022-0949. PMID 15767304. S2CID 18540594. Diarsipkan (PDF) dari versi aslinya tanggal 17 Oktober 2019. Diakses tanggal 31 Oktober 2019.
- ^ Skadhauge, E.; Erlwanger, K.H.; Ruziwa, S.D.; Dantzer, V.; Elbrønd, V.S.; Chamunorwa, J.P. (2003). "Does the ostrich (Struthio camelus) coprodeum have the electrophysiological properties and microstructure of other birds?". Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology. 134 (4): 749–755. doi:10.1016/S1095-6433(03)00006-0. ISSN 1095-6433. PMID 12814783.
- ^ Preest, M.R.; Beuchat, C.A. (1997). "Ammonia excretion by hummingbirds". Nature. 386 (6625): 561–562. Bibcode:1997Natur.386..561P. doi:10.1038/386561a0. ISSN 0028-0836. S2CID 4372695.
- ^ Mora, J.; Martuscelli, J.; Ortiz Pineda, J.; Soberon, G. (1965). "The Regulation of Urea-Biosynthesis Enzymes in Vertebrates". Biochemical Journal. 96 (1): 28–35. doi:10.1042/bj0960028. ISSN 0264-6021. PMC 1206904. PMID 14343146.
- ^ Packard, G.C. (1966). "The Influence of Ambient Temperature and Aridity on Modes of Reproduction and Excretion of Amniote Vertebrates". The American Naturalist. 100 (916): 667–682. Bibcode:1966ANat..100..667P. doi:10.1086/282459. ISSN 0003-0147. JSTOR 2459303. S2CID 85424175.
- ^ Balgooyen, T.G. (1971). "Pellet Regurgitation by Captive Sparrow Hawks (Falco sparverius)" (PDF). Condor. 73 (3): 382–385. doi:10.2307/1365774. JSTOR 1365774. Diarsipkan dari asli (PDF) tanggal 4 April 2019. Diakses tanggal 30 Oktober 2019.
- ^ Xu, X.; Li, F.; Wang, Y.; Sullivan, C.; Zhang, F.; Zhang, X.; Sullivan, C.; Wang, X.; Zheng, X. (2018). "Exceptional dinosaur fossils reveal early origin of avian-style digestion". Scientific Reports. 8 (1): 14217. Bibcode:2018NatSR...814217Z. doi:10.1038/s41598-018-32202-x. ISSN 2045-2322. PMC 6155034. PMID 30242170.
- ^ Russell, Dale A. (1997). "Intelligence". Dalam Kevin Padian; Philip J. Currie (ed.). Encyclopedia of dinosaurs. San Diego: Academic Press. hlm. 370–372. ISBN 978-0-12-226810-6.
- ^ Brusatte 2012, hlm. 83
- ^ Huxley, Thomas H. (1868). "On the Animals which are most nearly intermediate between Birds and Reptiles". The Annals and Magazine of Natural History. 4 (2). London: Taylor & Francis: 66–75. Diakses tanggal October 31, 2019.
- ^ Heilmann 1926
- ^ Osborn, Henry Fairfield (1924). "Three new Theropoda, Protoceratops zone, central Mongolia" (PDF). American Museum Novitates (144). New York: American Museum of Natural History: 1–12. ISSN 0003-0082. Diarsipkan (PDF) dari versi aslinya tanggal 2007-06-12.
- ^ Ostrom, John H. (1973). "The ancestry of birds". Nature. 242 (5393). London: Nature Research: 136. Bibcode:1973NPhS..242..136O. doi:10.1038/242136a0. ISSN 0028-0836. S2CID 29873831.
- ^ Padian 1986, hlm. 1–55, "Saurischian Monophyly and the Origin of Birds" by Jacques Gauthier.
- ^ Mayr, Gerald; Pohl, Burkhard; Peters, D. Stefan (2005). "A Well-Preserved Archaeopteryx Specimen with Theropod Features" (PDF). Science. 310 (5753). Washington, D.C.: American Association for the Advancement of Science: 1483–1486. Bibcode:2005Sci...310.1483M. doi:10.1126/science.1120331. ISSN 0036-8075. PMID 16322455. S2CID 28611454. Diarsipkan (PDF) dari versi aslinya tanggal December 25, 2022. Diakses tanggal January 3, 2023.
- ^ Martin, Larry D. (2006). "A basal archosaurian origin for birds". Acta Zoologica Sinica. 50 (6): 977–990. ISSN 1674-5507.
- ^ a b Feduccia, Alan (October 1, 2002). "Birds are Dinosaurs: Simple Answer to a Complex Problem". The Auk. 119 (4). Washington, D.C.: American Ornithologists' Union: 1187–1201. doi:10.1642/0004-8038(2002)119[1187:BADSAT]2.0.CO;2 (tidak aktif July 1, 2025). ISSN 0004-8038. JSTOR 4090252. S2CID 86096746. Diarsipkan dari versi aslinya tanggal July 29, 2020. Diakses tanggal November 3, 2019. Pemeliharaan CS1: DOI nonaktif per Juli 2025 (link)
- ^ a b Switek, Brian (July 2, 2012). "Rise of the fuzzy dinosaurs". News. Nature. London: Nature Research. doi:10.1038/nature.2012.10933. ISSN 0028-0836. S2CID 123219913. Diarsipkan dari versi aslinya tanggal April 13, 2020. Diakses tanggal January 1, 2019.
- ^ Godefroit, P.; Sinitsa, S.M.; Dhouailly, D.; Bolotsky, Y.L.; Sizov, A.V.; McNamara, M.E.; Benton, M.J.; Spagna, P. (2014). "A Jurassic ornithischian dinosaur from Siberia with both feathers and scales" (PDF). Science. 345 (6195): 451–455. Bibcode:2014Sci...345..451G. doi:10.1126/science.1253351. hdl:1983/a7ae6dfb-55bf-4ca4-bd8b-a5ea5f323103. PMID 25061209. S2CID 206556907. Diarsipkan dari asli (PDF) tanggal February 9, 2019. Diakses tanggal July 27, 2016.
- ^ Xu, Xing; Norell, Mark A.; Kuang, Xuewen; et al. (2004). "Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids" (PDF). Nature. 431 (7009). London: Nature Research: 680–684. Bibcode:2004Natur.431..680X. doi:10.1038/nature02855. ISSN 0028-0836. PMID 15470426. S2CID 4381777.
- ^ Göhlich, Ursula B.; Chiappe, Luis M. (2006). "A new carnivorous dinosaur from the Late Jurassic Solnhofen archipelago" (PDF). Nature. 440 (7082). London: Nature Research: 329–332. Bibcode:2006Natur.440..329G. doi:10.1038/nature04579. ISSN 0028-0836. PMID 16541071. S2CID 4427002. Diarsipkan dari asli (PDF) tanggal April 26, 2019. Diakses tanggal November 1, 2019.
- ^ Kellner, Alexander W. A.; Wang, Xiaolin; Tischlinger, Helmut; et al. (2010). "The soft tissue of Jeholopterus (Pterosauria, Anurognathidae, Batrachognathinae) and the structure of the pterosaur wing membrane". Proceedings of the Royal Society B. 277 (1679). London: Royal Society: 321–329. doi:10.1098/rspb.2009.0846. ISSN 0962-8452. PMC 2842671. PMID 19656798.
- ^ Mayr, G.; Pittman, M.; Saitta, E.; Kaye, T.G.; Vinther, J. (2016). "Structure and homology of Psittacosaurus tail bristles". Palaeontology. 59 (6): 793–802. Bibcode:2016Palgy..59..793M. doi:10.1111/pala.12257. hdl:1983/029c668f-08b9-45f6-a0c5-30ce9256e593. S2CID 89156313.
- ^ a b Benton, M.J.; Dhouailly, D.; Jiang, B.; McNamara, M. (2019). "The Early Origin of Feathers". Trends in Ecology & Evolution. 34 (9): 856–869. Bibcode:2019TEcoE..34..856B. doi:10.1016/j.tree.2019.04.018. hdl:10468/8068. PMID 31164250. S2CID 174811556.
- ^ Barrett, P.M.; Evans, D.C.; Campione, N.E. (2015). "Evolution of dinosaur epidermal structures". Biology Letters. 11 (6) 20150229. doi:10.1098/rsbl.2015.0229. PMC 4528472. PMID 26041865.
- ^ Alibardi, Lorenzo; Knapp, Loren W.; Sawyer, Roger H. (2006). "Beta-keratin localization in developing alligator scales and feathers in relation to the development and evolution of feathers". Journal of Submicroscopic Cytology and Pathology. 38 (2–3). Siena: Nuova Immagine Editrice: 175–192. ISSN 1122-9497. PMID 17784647.
- ^ Lingham-Soliar, Theagarten (December 2003). "The dinosaurian origin of feathers: perspectives from dolphin (Cetacea) collagen fibers". Naturwissenschaften. 90 (12). Berlin: Springer Science+Business Media: 563–567. Bibcode:2003NW.....90..563L. doi:10.1007/s00114-003-0483-7. ISSN 0028-1042. PMID 14676953. S2CID 43677545.
- ^ a b Feduccia, Alan; Lingham-Soliar, Theagarten; Hinchliffe, J. Richard (November 2005). "Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence". Journal of Morphology. 266 (2). Hoboken, NJ: John Wiley & Sons: 125–166. Bibcode:2005JMorp.266..125F. doi:10.1002/jmor.10382. ISSN 0362-2525. PMID 16217748. S2CID 15079072.
- ^ Lingham-Soliar, Theagarten; Feduccia, Alan; Wang, Xiaolin (2007). "A new Chinese specimen indicates that 'protofeathers' in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres". Proceedings of the Royal Society B. 274 (1620). London: Royal Society: 1823–1829. Bibcode:2007PBioS.274.1823L. doi:10.1098/rspb.2007.0352. ISSN 0962-8452. PMC 2270928. PMID 17521978.
- ^ Prum, Richard O. (2003). "Are Current Critiques Of The Theropod Origin Of Birds Science? Rebuttal To Feduccia 2002". The Auk. 120 (2). Washington, D.C.: American Ornithologists' Union: 550–561. doi:10.1642/0004-8038(2003)120[0550:ACCOTT]2.0.CO;2 (tidak aktif July 1, 2025). ISSN 0004-8038. JSTOR 4090212. Pemeliharaan CS1: DOI nonaktif per Juli 2025 (link)
- ^ Wellnhofer, Peter (1988). "A New Specimen of Archaeopteryx". Science. 240 (4860). Washington, D.C.: American Association for the Advancement of Science: 1790–1792. Bibcode:1988Sci...240.1790W. doi:10.1126/science.240.4860.1790. ISSN 0036-8075. JSTOR 1701652. PMID 17842432. S2CID 32015255.
- —— (1988). "Ein neuer Exemplar von Archaeopteryx". Archaeopteryx. 6: 1–30.
- ^ Schweitzer, Mary H.; Watt, J.A.; Avci, R.; et al. (1999). "Beta-keratin specific immunological reactivity in feather-like structures of the Cretaceous Alvarezsaurid, Shuvuuia deserti". Journal of Experimental Zoology Part B. 285 (2). Hoboken, NJ: Wiley-Blackwell: 146–157. Bibcode:1999JEZ...285..146S. doi:10.1002/(SICI)1097-010X(19990815)285:2<146::AID-JEZ7>3.0.CO;2-A. ISSN 1552-5007. PMID 10440726.
- ^ "Archaeopteryx: An Early Bird". Berkeley: University of California Museum of Paleontology. Diarsipkan dari versi aslinya tanggal 3 Februari 2021. Diakses tanggal 30 Oktober 2019.
- ^ O'Connor, Patrick M.; Claessens, Leon P. A. M. (2005). "Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs". Nature. 436 (7048). London: Nature Research: 253–256. Bibcode:2005Natur.436..253O. doi:10.1038/nature03716. ISSN 0028-0836. PMID 16015329. S2CID 4390587.
- ^ O'Connor, Patrick M.; Claessens, Leon P. A. M. (July 2005). "Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs". Nature (dalam bahasa Inggris). 436 (7048): 253–256. Bibcode:2005Natur.436..253O. doi:10.1038/nature03716. ISSN 0028-0836. PMID 16015329. Diarsipkan dari versi aslinya tanggal December 25, 2024. Diakses tanggal December 25, 2024.
- ^ "Meat-eating dinosaur from Argentina had bird-like breathing system". University of Michigan News. Ann Arbor, MI: Office of the Vice President for Communications; Regents of the University of Michigan. October 2, 2008. Diarsipkan dari versi aslinya tanggal July 30, 2020. Diakses tanggal November 2, 2019.
- ^ Xu, Xing; Norell, Mark A. (2004). "A new troodontid dinosaur from China with avian-like sleeping posture" (PDF). Nature. 431 (7010). London: Nature Research: 838–841. Bibcode:2004Natur.431..838X. doi:10.1038/nature02898. ISSN 0028-0836. PMID 15483610. S2CID 4362745.
- ^ Norell, Mark A.; Clark, James M.; Chiappe, Luis M.; et al. (1995). "A nesting dinosaur". Nature. 378 (6559). London: Nature Research: 774–776. Bibcode:1995Natur.378..774N. doi:10.1038/378774a0. ISSN 0028-0836. S2CID 4245228.
- ^ Varricchio, David J.; Moore, Jason R.; Erickson, Gregory M.; et al. (2008). "Avian Paternal Care Had Dinosaur Origin". Science. 322 (5909). Washington, D.C.: American Association for the Advancement of Science: 1826–1828. Bibcode:2008Sci...322.1826V. doi:10.1126/science.1163245. ISSN 0036-8075. PMID 19095938. S2CID 8718747.
- ^ Wings, Oliver (2007). "A review of gastrolith function with implications for fossil vertebrates and a revised classification" (PDF). Palaeontologica Polonica. 52 (1). Warsaw: Institute of Paleobiology, Polish Academy of Sciences: 1–16. ISSN 0567-7920. Diarsipkan (PDF) dari versi aslinya tanggal 2008-12-17. Diakses tanggal November 2, 2019.
- ^ Longrich, N.R.; Tokaryk, T.; Field, D.J. (2011). "Mass extinction of birds at the Cretaceous–Paleogene (K–Pg) boundary". Proceedings of the National Academy of Sciences. 108 (37): 15253–15257. Bibcode:2011PNAS..10815253L. doi:10.1073/pnas.1110395108. PMC 3174646. PMID 21914849.
- ^ a b Renne, P.R.; Deino, A.L.; Hilgen, F.J.; Kuiper, K.F.; Mark, D.F.; Mitchell, W.S.; Morgan, L.E.; Mundil, R.; Smit, J. (2013). "Time scales of critical events around the Cretaceous-Paleogene boundary". Science. 339 (6120): 684–687. Bibcode:2013Sci...339..684R. doi:10.1126/science.1230492. PMID 23393261. S2CID 6112274.
- ^ a b c d e Brusatte, S.L.; Butler, R.J.; Barrett, P.M.; Carrano, M.T.; Evans, D.C.; Lloyd, G.T.; Mannion, P.D.; Norell, M.A.; Peppe, D.J.; Upchurch, P.; Williamson, T.E. (2015). "The extinction of the dinosaurs". Biological Reviews. 90 (2): 628–642. doi:10.1111/brv.12128. hdl:20.500.11820/176e5907-26ec-4959-867f-0f2e52335f88. PMID 25065505. S2CID 115134484.
- ^ a b MacLeod, N.; Rawson, P.F.; Forey, P.L.; et al. (1997). "The Cretaceous–Tertiary biotic transition". Journal of the Geological Society. 154 (2): 265–292. Bibcode:1997JGSoc.154..265M. doi:10.1144/gsjgs.154.2.0265. ISSN 0016-7649. S2CID 129654916.
- ^ a b Archibald, J.D.; Clemens, W.A. (1982). "Late Cretaceous Extinctions". American Scientist. 70 (4): 377–385. Bibcode:1982AmSci..70..377A. JSTOR 27851545.
- ^ Jablonski, D. (1991). "Extinctions: a paleontological perspective". Science. 253 (5021): 754–757. Bibcode:1991Sci...253..754J. doi:10.1126/science.253.5021.754. PMID 17835491.
- ^ Longrich, N.R.; Bhullar, B.-A. S.; Gauthier, J.A. (2012). "Mass extinction of lizards and snakes at the Cretaceous–Paleogene boundary". Proceedings of the National Academy of Sciences. 109 (52): 21396–21401. Bibcode:2012PNAS..10921396L. doi:10.1073/pnas.1211526110. ISSN 0027-8424. PMC 3535637. PMID 23236177.
- ^ Field, D.J.; Bercovici, A.; Berv, J.S.; Dunn, R.; Fastovsky, D.E.; Lyson, T.R.; Vajda, V.; Gauthier, J.A. (2018). "Early evolution of modern birds structured by global forest collapse at the end-Cretaceous mass extinction". Current Biology. 28 (11): 1825–1831. Bibcode:2018CBio...28E1825F. doi:10.1016/j.cub.2018.04.062. PMID 29804807. S2CID 44075214.
- ^ a b Larson, D.W.; Brown, C.M.; Evans, D.C. (2016). "Dental disparity and ecological stability in bird-like dinosaurs prior to the end-Cretaceous mass extinction". Current Biology. 26 (10): 1325–1333. Bibcode:2016CBio...26.1325L. doi:10.1016/j.cub.2016.03.039. PMID 27112293. S2CID 3937001.
- ^ a b Le Loeuff, J. (2012). "Paleobiogeography and biodiversity of Late Maastrichtian dinosaurs: how many dinosaur species went extinct at the Cretaceous-Tertiary boundary?". Bulletin de la Société Géologique de France. 183 (6): 547–559. doi:10.2113/gssgfbull.183.6.547. ISSN 0037-9409.
- ^ Carpenter, K. (1983). "Evidence suggesting gradual extinction of latest Cretaceous dinosaurs". Naturwissenschaften. 70 (12): 611–612. Bibcode:1983NW.....70..611C. doi:10.1007/BF00377404. S2CID 20078285.
- ^ Russell, D.A. (1984). "The gradual decline of the dinosaurs—fact or fallacy?". Nature. 307 (5949): 360–361. Bibcode:1984Natur.307..360R. doi:10.1038/307360a0. S2CID 4269426.
- ^ Fastovsky, D.E.; Huang, Y.; Hsu, J.; Martin-McNaughton, J.; Sheehan, P.M.; Weishampel, D.B. (2004). "Shape of Mesozoic dinosaur richness" (PDF). Geology. 32 (10): 877–880. Bibcode:2004Geo....32..877F. doi:10.1130/G20695.1. Diarsipkan (PDF) dari versi aslinya tanggal 4 Februari 2023. Diakses tanggal 26 Januari 2023.
- ^ Sullivan, R.M. (2006). "The shape of Mesozoic dinosaur richness: a reassessment". Dalam Lucas, S.G.; Sullivan, R.M. (ed.). Late Cretaceous vertebrates from the Western Interior. New Mexico Museum of Natural History and Science Bulletin. Vol. 35. hlm. 403–405.
- ^ Chiarenza, A.A.; Mannion, P.D.; Lunt, D.J.; Farnsworth, A.; Jones, L.A.; Kelland, S.J.; Allison, P.A. (2019). "Ecological niche modelling does not support climatically-driven dinosaur diversity decline before the Cretaceous/Paleogene mass extinction". Nature Communications. 10 (1): 1–14. Bibcode:2019NatCo..10.1091C. doi:10.1038/s41467-019-08997-2. PMC 6403247. PMID 30842410.
- ^ Lloyd, G.T. (2012). "A refined modelling approach to assess the influence of sampling on palaeobiodiversity curves: new support for declining Cretaceous dinosaur richness". Biology Letters. 8 (1): 123–126. doi:10.1098/rsbl.2011.0210. PMC 3259943. PMID 21508029. S2CID 1376734.
- ^ Sakamoto, M.; Benton, M.J.; Venditti, C. (2016). "Dinosaurs in decline tens of millions of years before their final extinction". Proceedings of the National Academy of Sciences. 113 (18): 5036–5040. Bibcode:2016PNAS..113.5036S. doi:10.1073/pnas.1521478113. PMC 4983840. PMID 27092007.
- ^ Barrett, P.M.; McGowan, A.J.; Page, V. (2009). "Dinosaur diversity and the rock record". Proceedings of the Royal Society B: Biological Sciences. 276 (1667): 2667–2674. doi:10.1098/rspb.2009.0352. PMC 2686664. PMID 19403535.
- ^ Upchurch, P.; Mannion, P.D.; Benson, R.B.; Butler, R.J.; Carrano, M.T. (2011). "Geological and anthropogenic controls on the sampling of the terrestrial fossil record: a case study from the Dinosauria". Geological Society, London, Special Publications. 358 (1): 209–240. Bibcode:2011GSLSP.358..209U. doi:10.1144/SP358.14. S2CID 130777837.
- ^ Randall 2015
- ^ Alvarez, L.W.; Alvarez, W.; Asaro, F.; Michel, H.V. (1980). "Extraterrestrial Cause for the Cretaceous-Tertiary Extinction" (PDF). Science. 208 (4448): 1095–1108. Bibcode:1980Sci...208.1095A. CiteSeerX 10.1.1.126.8496. doi:10.1126/science.208.4448.1095. ISSN 0036-8075. PMID 17783054. S2CID 16017767. Diarsipkan dari asli (PDF) tanggal July 8, 2010. Diakses tanggal October 30, 2019.
- ^ Bohor, B.F.; Modreski, P.J.; Foord, E.E. (1987). "Shocked quartz in the Cretaceous-Tertiary boundary clays: Evidence for a global distribution". Science. 236 (4802): 705–709. Bibcode:1987Sci...236..705B. doi:10.1126/science.236.4802.705. PMID 17748309. S2CID 31383614. Diarsipkan dari versi aslinya tanggal 26 Februari 2021. Diakses tanggal 8 Februari 2021.
- ^ Hildebrand, A.R.; Penfield, G.T.; Kring, D.A.; Pilkington, M.; Camargo, Z.A.; Jacobsen, S.B.; Boynton, W.V. (1991). "Chicxulub crater: a possible Cretaceous/Tertiary boundary impact crater on the Yucatan Peninsula, Mexico". Geology. 19 (9): 867–871. Bibcode:1991Geo....19..867H. doi:10.1130/0091-7613(1991)019<0867:CCAPCT>2.3.CO;2.
- ^ Pope, K.O.; Ocampo, A.C.; Kinsland, G.L.; et al. (1996). "Surface expression of the Chicxulub crater". Geology. 24 (6). Boulder, CO: Geological Society of America: 527–530. Bibcode:1996Geo....24..527P. doi:10.1130/0091-7613(1996)024<0527:SEOTCC>2.3.CO;2. ISSN 0091-7613. PMID 11539331.
- ^ Schulte, P.; Alegret, L.; Arenillas, I.; Arz, J.A.; Barton, P.J.; Bown, P.R.; Bralower, T.J.; Christeson, G.L.; Claeys, P.; Cockell, C.S.; Collins, G.S.; Deutsch, A.; Goldin, T.J.; Goto, K.; Grajales-Nishimura, J.M.; Grieve, R.A.F.; Gulick , S.P.S.; Johnson, K.R.; Kiessling, W.; Koeberl, C.; Kring, D.A.; MacLeod, K.G.; Matsui, T.; Melosh, J.; Montanari, A.; Morgan, J.V.; Neal, C.R.; Nichols, D.J.; Norris, R.D.; Pierazzo, E.; Ravizza, G.; Rebolledo-Vieyra, M.; Uwe Reimold, W.; Robin, E.; Salge, T.; Speijer, R.P.; Sweet, A.R.; Urrutia-Fucugauchi, J.; Vajda, V.; Whalen, M.T.; Willumsen, P.S. (2010). "The Chicxulub asteroid impact and mass extinction at the Cretaceous-Paleogene boundary". Science. 327 (5970): 1214–1218. Bibcode:2010Sci...327.1214S. doi:10.1126/science.1177265. PMID 20203042. S2CID 2659741.
- ^ Kring, D. A. (2007). "The Chicxulub impact event and its environmental consequences at the Cretaceous–Tertiary boundary". Palaeogeography, Palaeoclimatology, Palaeoecology. 255 (1–2): 4–21. Bibcode:2007PPP...255....4K. doi:10.1016/j.palaeo.2007.02.037.
- ^ a b Chiarenza, A.A.; Farnsworth, A.; Mannion, P.D.; Lunt, D.J.; Valdes, P.J.; Morgan, J.V.; Allison, P.A. (2020). "Asteroid impact, not volcanism, caused the end-Cretaceous dinosaur extinction". Proceedings of the National Academy of Sciences. 117 (29): 17084–17093. Bibcode:2020PNAS..11717084C. doi:10.1073/pnas.2006087117. PMC 7382232. PMID 32601204.
- ^ Ivanov, B.A. (2005). "Numerical Modeling of the Largest Terrestrial Meteorite Craters". Solar System Research. 39 (5): 381–409. Bibcode:2005SoSyR..39..381I. doi:10.1007/s11208-005-0051-0. S2CID 120305483.
- ^ Matsui, T.; Imamura, F.; Tajika, E.; Nakano, Y.; Fujisawa, Y. (2002). "Generation and propagation of a tsunami from the Cretaceous-Tertiary impact event". Geological Society of America Special Papers. 356: 69–78. doi:10.1130/0-8137-2356-6.69. ISBN 978-0-8137-2356-3.
- ^ Robertson, D.S.; McKenna, M.C.; Toon, O.B.; et al. (2004). "Survival in the first hours of the Cenozoic" (PDF). Geological Society of America Bulletin. 116 (5–6): 760–768. Bibcode:2004GSAB..116..760R. doi:10.1130/B25402.1. ISSN 0016-7606. Diarsipkan dari asli (PDF) tanggal 18 September 2012. Diakses tanggal 15 Juni 2011.
- ^ Robertson, D.S.; Lewis, W.M.; Sheehan, P.M.; Toon, O.B. (2013). "K-Pg extinction: Reevaluation of the heat-fire hypothesis". Journal of Geophysical Research: Biogeosciences. 118 (1): 329–336. Bibcode:2013JGRG..118..329R. doi:10.1002/jgrg.20018. S2CID 17015462.
- ^ Pope, K.O.; Baines, K.H.; Ocampo, A.C.; Ivanov, B.A. (1997). "Energy, volatile production, and climatic effects of the Chicxulub Cretaceous/Tertiary impact". Journal of Geophysical Research: Planets. 102 (E9): 21645–21664. Bibcode:1997JGR...10221645P. doi:10.1029/97JE01743. PMID 11541145. S2CID 8447773.
- ^ a b Ohno, S.; Kadono, T.; Kurosawa, K.; Hamura, T.; Sakaiya, T.; Shigemori, K.; Hironaka, Y.; Sano, T.; Watari, T.; Otani, K.; Matsui, T.; Sugita, S. (2014). "Production of sulphate-rich vapour during the Chicxulub impact and implications for ocean acidification". Nature Geoscience. 7 (4): 279–282. Bibcode:2014NatGe...7..279O. doi:10.1038/ngeo2095.
- ^ Kaiho, K.; Oshima, N.; Adachi, K.; Adachi, Y.; Mizukami, T.; Fujibayashi, M.; Saito, R. (2016). "Global climate change driven by soot at the K-Pg boundary as the cause of the mass extinction". Scientific Reports. 6 (1): 1–13. Bibcode:2016NatSR...628427K. doi:10.1038/srep28427. PMC 4944614. PMID 27414998.
- ^ Lyons, S.L.; Karp, A.T.; Bralower, T.J.; Grice, K.; Schaefer, B.; Gulick, S.P.; Morgan, J.V.; Freeman, K.H. (2020). "Organic matter from the Chicxulub crater exacerbated the K–Pg impact winter". Proceedings of the National Academy of Sciences. 117 (41): 25327–25334. Bibcode:2020PNAS..11725327L. doi:10.1073/pnas.2004596117. PMC 7568312. PMID 32989138.
- ^ Chenet, A.L.; Courtillot, V.; Fluteau, F.; Gérard, M.; Quidelleur, X.; Khadri, S.F.R.; Subbarao, K.V.; Thordarson, T. (2009). "Determination of rapid Deccan eruptions across the Cretaceous-Tertiary boundary using paleomagnetic secular variation: 2. Constraints from analysis of eight new sections and synthesis for a 3500-m-thick composite section" (PDF). Journal of Geophysical Research: Solid Earth. 114 (B6): B06103. Bibcode:2009JGRB..114.6103C. doi:10.1029/2008JB005644. S2CID 140541003.
- ^ Schoene, B.; Eddy, M.P.; Samperton, K.M.; Keller, C.B.; Keller, G.; Adatte, T.; Khadri, S.F. (2019). "U-Pb constraints on pulsed eruption of the Deccan Traps across the end-Cretaceous mass extinction". Science. 363 (6429): 862–866. Bibcode:2019Sci...363..862S. doi:10.1126/science.aau2422. OSTI 1497969. PMID 30792300. S2CID 67876950.
- ^ a b McLean, D.M. (1985). "Deccan Traps mantle degassing in the terminal Cretaceous marine extinctions". Cretaceous Research. 6 (3): 235–259. Bibcode:1985CrRes...6..235M. doi:10.1016/0195-6671(85)90048-5.
- ^ Self, S.; Widdowson, M.; Thordarson, T.; Jay, A.E. (2006). "Volatile fluxes during flood basalt eruptions and potential effects on the global environment: A Deccan perspective". Earth and Planetary Science Letters. 248 (1–2): 518–532. Bibcode:2006E&PSL.248..518S. doi:10.1016/j.epsl.2006.05.041.
- ^ Tobin, T.S.; Bitz, C.M.; Archer, D. (2017). "Modeling climatic effects of carbon dioxide emissions from Deccan Traps volcanic eruptions around the Cretaceous–Paleogene boundary". Palaeogeography, Palaeoclimatology, Palaeoecology. 478: 139–148. Bibcode:2017PPP...478..139T. doi:10.1016/j.palaeo.2016.05.028.
- ^ Schmidt, A.; Skeffington, R.A.; Thordarson, T.; Self, S.; Forster, P.M.; Rap, A.; Ridgwell, A.; Fowler, D.; Wilson, M.; Mann, G.W.; Wignall, P.B.; Carslaw, K.S. (2016). "Selective environmental stress from sulphur emitted by continental flood basalt eruptions" (PDF). Nature Geoscience. 9 (1): 77–82. Bibcode:2016NatGe...9...77S. doi:10.1038/ngeo2588. S2CID 59518452. Diarsipkan (PDF) dari versi aslinya tanggal 22 September 2017.
- ^ Hofman, C.; Féraud, G.; Courtillot, V. (2000). "40Ar/39Ar dating of mineral separates and whole rocks from the Western Ghats lava pile: further constraints on duration and age of the Deccan traps". Earth and Planetary Science Letters. 180 (1–2): 13–27. Bibcode:2000E&PSL.180...13H. doi:10.1016/S0012-821X(00)00159-X. ISSN 0012-821X.
- ^ Sahni, A. (1988). "Cretaceous-Tertiary boundary events: Mass extinctions, iridium enrichment and Deccan volcanism". Current Science. 57 (10): 513–519. JSTOR 24090754.
- ^ Glasby, G.P.; Kunzendorf, H. (1996). "Multiple factors in the origin of the Cretaceous/Tertiary boundary: the role of environmental stress and Deccan Trap volcanism". Geologische Rundschau. 85 (2): 191–210. Bibcode:1996IJEaS..85..191G. doi:10.1007/BF02422228. PMID 11543126. S2CID 19155384.
- ^ Alvarez, L.W. (1987). Mass Extinctions Caused by Large Bolide Impacts (Report). Laboratorium Nasional Lawrence Berkeley. hlm. 39. LBL-22786. Diakses tanggal 27 Januari 2021.
- ^ Alvarez 1997, hlm. 130–146, chpt. 7: "The World after Chicxulub".
- ^ Renne, P.R.; Sprain, C.J.; Richards, M.A.; Self, S.; Vanderkluysen, L.; Pande, K. (2015). "State shift in Deccan volcanism at the Cretaceous-Paleogene boundary, possibly induced by impact". Science. 350 (6256): 76–78. Bibcode:2015Sci...350...76R. doi:10.1126/science.aac7549. PMID 26430116. S2CID 30612906. Diarsipkan dari versi aslinya tanggal 23 Maret 2023. Diakses tanggal 28 Oktober 2022.
- ^ Richards, M.A.; Alvarez, W.; Self, S.; Karlstrom, L.; Renne, P.R.; Manga, M.; Sprain, C.J.; Smit, J.; Vanderkluysen, L.; Gibson, S.A. (2015). "Triggering of the largest Deccan eruptions by the Chicxulub impact". Geological Society of America Bulletin. 127 (11–12): 1507–1520. Bibcode:2015GSAB..127.1507R. doi:10.1130/B31167.1. hdl:1871.1/cc9361fe-f586-44a0-90ac-f5c513e9920b. OSTI 1512141. S2CID 3463018. Diarsipkan dari versi aslinya tanggal 22 Oktober 2020. Diakses tanggal 8 Februari 2021.
- ^ Khazins, V.; Shuvalov, V. (2019). "Chicxulub Impact as a Trigger of One of Deccan Volcanism Phases: Threshold of Seismic Energy Density". Dalam Kocharyan, G.; Lyakhov, A. (ed.). Trigger Effects in Geosystems. Springer Proceedings in Earth and Environmental Sciences. Cham: Springer. hlm. 523–530. doi:10.1007/978-3-030-31970-0_55. ISBN 978-3-030-31969-4. S2CID 210277965.
- ^ Archibald, J.D.; Clemens, W.A.; Padian, K.; Rowe, T.; Macleod, N.; Barrett, P.M.; Gale, A.; Holroyd, P.; Sues, H.-D.; Arens, N.C.; Horner, J.R.; Wilson, G.P.; Goodwin, M.B.; Brochu, C.A.; Lofgren, D.L.; Hurlbert, S.H.; Hartman, J.H.; Eberth, D.A.; Wignall, P.B.; Currie, P.J.; Weil, A.; Prasad, G.V.R.; Dingus, L.; Courtillot, V.; Milner, A.; Milner, A.; Bajpai, S.; Ward, D.J.; Sahni, A. (2010). "Cretaceous extinctions: multiple causes". Science. 328 (5981): 973, author reply 975–6. doi:10.1126/science.328.5981.973-a. PMID 20489004.
- ^ Courtillot, V.; Fluteau, F. (2010). "Cretaceous extinctions: the volcanic hypothesis". Science. 328 (5981): 973–974. Bibcode:2010Sci...328..973C. doi:10.1126/science.328.5981.973-b. PMID 20489003.
- ^ Keller, G. (2014). "Deccan volcanism, the Chicxulub impact, and the end-Cretaceous mass extinction: Coincidence? Cause and effect". Geological Society of America Special Papers. 505: 57–89. doi:10.1130/2014.2505(03). ISBN 978-0-8137-2505-5.
- ^ Schulte, P.; Alegret, L.; Arenillas, I.; Arz, J.A.; Barton, P.J.; Bown, P.R.; Bralower, T.J.; Christeson, G.L.; Claeys, P.; Cockell, C.S.; Collins, G.S.; Deutsch, A.; Goldin, T.J.; Goto, K.; Grajales-Nishimura, J.M.; Grieve, R.A.F.; Gulick , S.P.S.; Johnson, K.R.; Kiessling, W.; Koeberl, C.; Kring, D.A.; MacLeod, K.G.; Matsui, T.; Melosh, J.; Montanari, A.; Morgan, J.V.; Neal, C.R.; Nichols, D.J.; Norris, R.D.; Pierazzo, E.; Ravizza, G.; Rebolledo-Vieyra, M.; Uwe Reimold, W.; Robin, E.; Salge, T.; Speijer, R.P.; Sweet, A.R.; Urrutia-Fucugauchi, J.; Vajda, V.; Whalen, M.T.; Willumsen, P.S. (2010). "Response—Cretaceous extinctions". Science. 328 (5981): 975–976. Bibcode:2010Sci...328..975S. doi:10.1126/science.328.5981.975.
- ^ Fassett, J.E.; Heaman, L.M.; Simonetti, A. (2011). "Direct U–Pb dating of Cretaceous and Paleocene dinosaur bones, San Juan Basin, New Mexico". Geology. 39 (2): 159–162. Bibcode:2011Geo....39..159F. doi:10.1130/G31466.1. ISSN 0091-7613.
- ^ Fassett, J.E.; Heaman, L.M.; Simonetti, A. (2009). "New geochronologic and stratigraphic evidence confirms the Paleocene age of the dinosaur-bearing Ojo Alamo Sandstone and Animas Formation in the San Juan Basin, New Mexico and Colorado". Palaeontologia Electronica. 12 (1): 3A. Diarsipkan dari versi aslinya tanggal March 18, 2021. Diakses tanggal January 27, 2021.
- ^ Sloan, R.E.; Rigby, J.K. Jr.; Van Valen, L.M.; et al. (1986). "Gradual Dinosaur Extinction and Simultaneous Ungulate Radiation in the Hell Creek Formation". Science. 232 (4750): 629–633. Bibcode:1986Sci...232..629S. doi:10.1126/science.232.4750.629. ISSN 0036-8075. PMID 17781415. S2CID 31638639.
- ^ Lucas, S.G.; Sullivan, R.M.; Cather, S.M.; Jasinski, S.E.; Fowler, D.W.; Heckert, A.B.; Spielmann, J.A.; Hunt, A.P. (2009). "No definitive evidence of Paleocene dinosaurs in the San Juan Basin". Palaeontologia Electronica. 12 (2): 8A.
- ^ Renne, P.R.; Goodwin, M.B. (2012). "Direct U-Pb dating of Cretaceous and Paleocene dinosaur bones, San Juan Basin, New Mexico: COMMENT". Geology. 40 (4): e259. Bibcode:2012Geo....40E.259R. doi:10.1130/G32521C.1.
- ^ Lofgren, D.L.; Hotton, C.L.; Runkel, A.C. (1990). "Reworking of Cretaceous dinosaurs into Paleocene channel, deposits, upper Hell Creek Formation, Montana". Geology. 18 (9): 874–877. Bibcode:1990Geo....18..874L. doi:10.1130/0091-7613(1990)018<0874:ROCDIP>2.3.CO;2.
- ^ Koenig, A.E.; Lucas, S.G.; Neymark, L.A.; Heckert, A.B.; Sullivan, R.M.; Jasinski, S.E.; Fowler, D.W. (2012). "Direct U-Pb dating of Cretaceous and Paleocene dinosaur bones, San Juan Basin, New Mexico: COMMENT". Geology. 40 (4): e262. Bibcode:2012Geo....40E.262K. doi:10.1130/G32154C.1.
- ^ "Dinosaur". Merriam-Webster Dictionary. Diakses tanggal November 7, 2019.
- ^ Sarjeant 1995, hlm. 255–284, chpt. 15: "The Dinosaurs and Dinomania over 150 Years" by Hugh S. Torrens.
- ^ Currie & Padian 1997, hlm. 347–350, "History of Dinosaur Discoveries: First Golden Period" by Brent H. Breithaupt.
- ^ Dickens 1853, hlm. 1, chpt. I: "London. Michaelmas Term lately over, and the Lord Chancellor sitting in Lincoln's Inn Hall. Implacable November weather. As much mud in the streets, as if the waters had but newly retired from the face of the earth, and it would not be wonderful to meet a Megalosaurus, forty feet long or so, waddling like an elephantine lizard up Holborn Hill."
- ^ Farlow & Brett-Surman 1997, hlm. 675–697, chpt. 43: "Dinosaurs and the Media" by Donald F. Glut and M.K. Brett-Surman.
- ^ Lee, Newton; Madej, Krystina (2012). "Early Animation: Gags and Situations". Disney Stories. hlm. 17–24. doi:10.1007/978-1-4614-2101-6_3. ISBN 978-1-4614-2100-9. S2CID 192335675.
Bibliografi
sunting- Alvarez, Walter (1997). T. rex and the Crater of Doom. Princeton, NJ: Princeton University Press. ISBN 978-0-691-01630-6. LCCN 96049208. OCLC 1007846558. Diakses tanggal November 4, 2019.
- Bakker, Robert T. (1986). The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and Their Extinction. New York: William Morrow and Company. ISBN 978-0-688-04287-5. LCCN 86012643. OCLC 13699558. Diakses tanggal November 6, 2019.
- Benton, Michael J. (2015). Vertebrate Palaeontology (Edisi 4th [completely revised]). Oxford: Wiley Blackwell. ISBN 978-1-118-40684-7.
- Benton, Michael J. (2005). Vertebrate Palaeontology (Edisi 3rd). Malden, MA: Blackwell Publishing. ISBN 978-0-632-05637-8. LCCN 2003028152. OCLC 53970617. Diakses tanggal October 30, 2019.
- Brusatte, Stephen L. (2012). Benton, Michael J. (ed.). Dinosaur Paleobiology. Topics in Paleobiology. Foreword by Michael J. Benton. Hoboken, NJ: Wiley-Blackwell. Bibcode:2012dipa.book.....B. doi:10.1002/9781118274071. ISBN 978-0-470-65658-7. LCCN 2011050466. OCLC 781864955.
- Chiappe, Luis M.; Witmer, Lawrence M., ed. (2002). Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press. ISBN 978-0-520-20094-4. LCCN 2001044600. OCLC 901747962.
- Colbert, Edwin H. (1971) [Originally published, New York: E. P. Dutton, 1968; London: Evans Brothers Ltd, 1969]. Men and Dinosaurs: The Search in Field and Laboratory. Harmondsworth: Penguin. ISBN 978-0-14-021288-4. OCLC 16208760. Diakses tanggal October 31, 2019.
- Cowen, Richard (2005). History of Life (Edisi 4th). Malden, MA: Blackwell Publishing. ISBN 978-1-4051-1756-2. LCCN 2003027993. OCLC 53970577. The 5th edition of the book is available from the Internet Archive. Retrieved 2019-10-19.
- Currie, Philip J.; Padian, Kevin, ed. (1997). Encyclopedia of Dinosaurs. San Diego, CA: Academic Press. ISBN 978-0-12-226810-6. LCCN 97023430. OCLC 436848919. Diakses tanggal October 30, 2019.
- Currie, Philip J.; Koppelhus, Eva B.; Shugar, Martin A.; Wright, Joanna L., ed. (2004). Feathered Dragons: Studies on the Transition from Dinosaurs to Birds. Life of the Past. Bloomington, IN: Indiana University Press. ISBN 978-0-253-34373-4. LCCN 2003019035. OCLC 52942941.
- Curry Rogers, Kristina A.; Wilson, Jeffrey A., ed. (2005). The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press. ISBN 978-0-520-24623-2. LCCN 2005010624. OCLC 879179542.
- Desmond, Adrian J. (1975). The Hot-Blooded Dinosaurs: A Revolution in Palaeontology. London: Blond & Briggs. ISBN 978-0-8037-3755-6. LCCN 76359907. OL 4933052M. Diakses tanggal October 30, 2019.
- Dickens, Charles (1853). Bleak House. London: Bradbury and Evans. Diakses tanggal November 7, 2019.
- Dodson, Peter; Gingerich, Philip D., ed. (1993). Functional Morphology and Evolution. American Journal of Science. Vol. 293A. New Haven, CT: Kline Geology Laboratory, Yale University. ISSN 0002-9599. OCLC 27781160.
- Dong, Zhiming (1992). Dinosaurian Faunas of China (Edisi English). Beijing; Berlin; New York: China Ocean Press; Springer-Verlag. ISBN 978-3-540-52084-9. LCCN 92207835. OCLC 26522845.
- Dyke, Gareth; Kaiser, Gary, ed. (2011). Living Dinosaurs: The Evolutionary History of Modern Birds. Chichester; Hoboken, NJ: Wiley-Blackwell. ISBN 978-0-470-65666-2. LCCN 2010043277. OCLC 729724640.
- Farlow, James O.; Brett-Surman, M.K., ed. (1997). The Complete Dinosaur. Bloomington, IN: Indiana University Press. ISBN 978-0-253-33349-0. LCCN 97-23698. OCLC 924985811. Diakses tanggal October 14, 2019.
- Foster, John R.; Lucas, Spencer G., ed. (2006). "Paleontology and Geology of the Upper Jurassic Morrison Formation". Bulletin of the New Mexico Museum of Natural History and Science. New Mexico Museum of Natural History and Science Bulletin. 36. Albuquerque, NM: New Mexico Museum of Natural History and Science. ISSN 1524-4156. OCLC 77520577. Diakses tanggal October 21, 2019.
- Glut, Donald F. (1997). Dinosaurs: The Encyclopedia. Foreword by Michael K. Brett-Surman. Jefferson, NC: McFarland & Company. ISBN 978-0-89950-917-4. LCCN 95047668. OCLC 33665881.
- Gunther, Robert Theodore, ed. (1968) [First printed in Oxford 1945]. Life and Letters of Edward Lhwyd. Early Science in Oxford. Vol. XIV. Preface by Albert Everard Gunther (Edisi Reprint). London: Dawsons of Pall Mall. ISBN 978-0-7129-0292-2. LCCN 22005926. OCLC 43529321. Diakses tanggal November 4, 2019.
- Hansell, Mike (2000). Bird Nests and Construction Behaviour. Pen and ink illustration by Raith Overhill. Cambridge: University of Cambridge Press. ISBN 978-0-521-46038-5. LCCN 99087681. OCLC 876286627. Diakses tanggal October 30, 2019.
- Heilmann, Gerhard (1926). The Origin of Birds. London; New York: H. F. & G. Witherby; D. Appleton & Company. LCCN 27001127. OCLC 606021642.
- Holmes, Thom (1998). Fossil Feud: The Rivalry of the First American Dinosaur Hunters. Parsippany, NJ: Julian Messner. ISBN 978-0-382-39149-1. LCCN 96013610. OCLC 34472600.
- Holtz, Thomas R. Jr. (2007). Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. Illustrated by Luis V. Rey. New York: Random House. ISBN 978-0-375-82419-7. LCCN 2006102491. OCLC 77486015. Diakses tanggal October 22, 2019.
- Lambert, David; The Diagram Group (1990). The Dinosaur Data Book: The Definitive, Fully Illustrated Encyclopedia of Dinosaurs. New York: Avon Books. ISBN 978-0-380-75896-8. LCCN 89092487. OCLC 21833417. Diakses tanggal October 14, 2019.
- Lessem, Don; Glut, Donald F. (1993). The Dinosaur Society's Dinosaur Encyclopedia. Illustrations by Tracy Lee Ford; scientific advisors, Peter Dodson, et al. New York: Random House. ISBN 978-0-679-41770-5. LCCN 94117716. OCLC 30361459. Diakses tanggal October 30, 2019.
- Lhuyd, Edward (1699). Lithophylacii Britannici ichnographia [British figured stones]. London: Ex Officina M.C. Diarsipkan dari asli tanggal August 14, 2020. Diakses tanggal November 4, 2019.
- Mayr, Gerald (2009). Paleogene Fossil Birds. Berlin: Springer-Verlag. doi:10.1007/978-3-540-89628-9. ISBN 978-3-540-89627-2. LCCN 2008940962. OCLC 916182693. S2CID 88941254. Diakses tanggal October 30, 2019.
- Norell, Mark; Gaffney, Eugene S.; Dingus, Lowell (2000) [Originally published as Discovering Dinosaurs in the American Museum of Natural History. New York: Knopf, 1995]. Discovering Dinosaurs: Evolution, Extinction, and the Lessons of Prehistory (Edisi Revised). Berkeley: University of California Press. ISBN 978-0-520-22501-5. LCCN 99053335. OCLC 977125867. Diakses tanggal October 30, 2019.
- Olshevsky, George (2000). An Annotated Checklist of Dinosaur Species by Continent. Mesozoic Meanderings. Vol. 3. Illustrated by Tracy Lee Ford. San Diego, CA: Publications Requiring Research. ISSN 0271-9428. LCCN 00708700. OCLC 44433611.
- Owen, Richard (1842). "Report on British Fossil Reptiles. Part II". Report of the Eleventh Meeting of the British Association for the Advancement of Science; Held at Plymouth in July 1841. London: John Murray. hlm. 60–204. LCCN 99030427. OCLC 1015526268. Diakses tanggal October 13, 2019.
- Padian, Kevin, ed. (1986). The Origin of Birds and the Evolution of Flight. Memoirs of the California Academy of Sciences. Vol. 8. San Francisco, CA: California Academy of Sciences. ISBN 978-0-940228-14-6. OCLC 946083441. OL 9826926M.
- Parsons, Keith M. (2001). Drawing out Leviathan: Dinosaurs and the Science Wars. Life in the Past. Bloomington, IN: Indiana University Press. ISBN 978-0-253-33937-9. LCCN 2001016803. OCLC 50174737. Diakses tanggal October 30, 2019.
- Paul, Gregory S. (1988). Predatory Dinosaurs of the World: A Complete Illustrated Guide. New York: Simon & Schuster. ISBN 978-0-671-61946-6. LCCN 88023052. OCLC 859819093. Diakses tanggal October 30, 2019.
- Paul, Gregory S., ed. (2000). The Scientific American Book of Dinosaurs (Edisi 1st). New York: St. Martin's Press. ISBN 978-0-312-26226-6. LCCN 2001269051. OCLC 45256074.
- Paul, Gregory S. (2010). The Princeton Field Guide to Dinosaurs. Princeton Field Guides. Princeton, NJ: Princeton University Press. ISBN 978-0-691-13720-9. LCCN 2010014916. OCLC 907619291.
- Plot, Robert (1677). The Natural History of Oxford-shire: Being an Essay toward the Natural History of England. Oxford; London: S. Millers. LCCN 11004267. OCLC 933062622. Diakses tanggal November 13, 2019.
- Randall, Lisa (2015). Dark Matter and the Dinosaurs: The Astounding Interconnectedness of the Universe. New York: HarperCollins. ISBN 978-0-06-232847-2. LCCN 2016427646. OCLC 962371431.
- Rupke, Nicolaas A. (1994). Richard Owen: Victorian Naturalist. New Haven: Yale University Press. ISBN 978-0-300-05820-8. LCCN 93005739. OCLC 844183804. Diakses tanggal November 5, 2019.
- Sarjeant, William A.S., ed. (1995). Vertebrate Fossils and the Evolution of Scientific Concepts: Writings in Tribute to Beverly Halstead, by Some of His Many Friends. Modern Geology, vol. 18. Amsterdam: Gordon and Breach Publishers. ISBN 978-2-88124-996-9. ISSN 0026-7775. LCCN 00500382. OCLC 34672546. "Reprint of papers published in a special volume of Modern geology [v. 18 (Halstead memorial volume), 1993], with five additional contributions.--Pref."
- Tanner, Lawrence H.; Spielmann, Justin A.; Lucas, Spencer G., ed. (2013). "The Triassic System: New Developments in Stratigraphy and Paleontology". Bulletin of the New Mexico Museum of Natural History and Science. New Mexico Museum of Natural History and Science Bulletin. 61. Albuquerque, NM: New Mexico Museum of Natural History and Science. ISSN 1524-4156. OCLC 852432407. Diakses tanggal October 21, 2019.
- Weishampel, David B.; Dodson, Peter; Osmólska, Halszka, ed. (2004). The Dinosauria (Edisi 2nd). Berkeley: University of California Press. ISBN 978-0-520-25408-4. LCCN 2004049804. OCLC 154697781.
Bacaan lanjutan
sunting| Sumber pustaka mengenai Dinosaurs |
- University of Southampton (September 29, 2021). "Two New Species of Large Predatory Dinosaur With Crocodile-Like Skulls Discovered on Isle of Wight". SciTechDaily.
- Zhou, Zhonghe (October 2004). "The origin and early evolution of birds: discoveries, disputes, and perspectives from fossil evidence" (PDF). Naturwissenschaften. 91 (10). Berlin: Springer Science+Business Media: 455–471. Bibcode:2004NW.....91..455Z. doi:10.1007/s00114-004-0570-4. ISSN 0028-1042. PMID 15365634. S2CID 3329625. Diarsipkan dari asli (PDF) tanggal July 21, 2011. Diakses tanggal November 6, 2019.
- Paul, Gregory S. (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore; London: Johns Hopkins University Press. ISBN 978-0-8018-6763-7. LCCN 2001000242. OCLC 1088130487..
- Stewart, Tabori & Chang (1997). The Humongous Book of Dinosaurs. New York: Stewart, Tabori & Chang. ISBN 978-1-55670-596-0. LCCN 97000398. OCLC 1037269801.
- Sternberg, Charles Mortram (1966) [Original edition published by E. Cloutier, printer to the King, 1946]. Canadian Dinosaurs. Geological Series. Vol. 54 (Edisi 2nd). Ottawa: National Museum of Canada. LCCN gs46000214. OCLC 1032865683.
